In
chapter four of his controversial book, Darwin's Black Box, Michael
Behe presents the argument that Rube Goldberg machines exist in living things
and that such machines are "irreducibly complex."Behe presents the argument that the existence of such machines cannot be
explained by mindless naturalistic mechanisms and are thus examples of
deliberate design.After all,
anyone who has watched cartoons as a child knows what a Rube Goldberg machine is
and that this machine will not work if any one part is removed.
So, how can something evolve in a stepwise way where each step is functionally
beneficial if there is no function until all the parts are in place?
As an example, consider the following scenario where Behe
describes a popular cartoon about the loud-mouthed rooster Foghorn Leghorn.
"...Foghorn would be walking along, notice a dollar bill or some other bait on the
ground, and pick it up.The dollar
was tied by a string to a stick that was propped against a ball.When the dollar bill was moved, the attached string pulled down the
stick, and the ball would start to roll away as Foghorn stared slack-jawed at
the developing action.The ball
then would fall of a cliff onto the raised end of a seesaw, smacking it down and
sending a rock with an attached piece of sandpaper hurtling into the air.On its upward journey the sandpaper would strike a match sticking out of
the cliff, which lit the fuse to a cannon.The cannon would fire; on its downward track the cannonball would hit the
rim of a funnel (the only allowance for error in the whole scenario), roll
around the edge a few times, and fall through.As it came out of the funnel, the cannonball would hit against a lever
that started a circular saw.The
saw would cut through a rope, which was holding up a telephone pole.Slowly the telephone pole would begin to fall, and too late Foghorn
Leghorn would realize that the fascinating show was at his expense.As he turns to run, the very tip of the telephone pole smacks him on the
head and drives him like a peg into the ground."1
Behe
goes on to say that this Foghorn Leghorn cartoon contraption, as a Rube
Goldberg-like machine, is "irreducibly complex." This of course means
that if any one part is removed, the whole machine fails and the desired end
result or function, does not occur.Behe
compares such thought contraptions to real life systems of functions within
living things, such as the clotting cascade in the blood coagulation pathway.This clotting cascade works in just about the same way that Foghorn
Leghorn was whacked into the ground by the telephone pole.Each event in the clotting cascade must happen before the next event can
occur.If any one event is blocked,
clotting will not occur at all.Behe
wonders how such a system could have evolved by mindless naturalistic
mechanisms?
Well,
we must first ask if Rube Goldberg machines really are "irreducibly
complex"?It is true that if one
part is removed the cascade is interrupted at that point. It is kind of
like taking a domino out of a line of dominos that are balanced on their ends.
The dominos after this point will not be knocked over by the previously
cascading dominos. In this way, the final
function of the cascade is rendered useless. However, the cascade is in
fact still able to function from the point of interference onwards. So,
its function cannot be said to be completely destroyed just because of a single
point of interruption. Also, by the very nature of cascades, more
parts can be added on the originating end of the cascade to make it longer and
longer still... like adding more and more dominos onto the end of a chain of
dominos. At first one starts with one domino, then two, three, four and so
on until one has a very long cascade set up.
Consider
the Foghorn Leghorn cascade again from this perspective. What if
the entire cascade describing Foghorn Leghorn’s demise started simply?Foghorn picks up the dollar that is attached directly to the telephone
pole by a string. When the string is
pulled, the carefully balanced telephone pole falls over and drives Foghorn into
the ground like a tent peg. Granted, this certainly is not nearly
as interesting or entertaining.But,
it would work - right?Now, what if
we add just one little part to the cascade?
Lets add the rope that holds up the telephone pole and a saw that cuts the rope.
The string is attached to the switch on the saw. When pulled, the string
turns the saw on and it cuts the rope and the pole falls. A bit more
interesting and it still works.Now
lets add one more little part.Lets
add the cannon. The string pulls a match and ignites the cannon and the cannon
ball hits the saw switch which cuts the rope that holds the pole… and now we
are getting a lot more interesting!We
are evolving a complex cascade one small part at a time - right?It sure looks that way.
This
argument is in fact commonly used as an explanation for the origins of such
apparently complex cascades as occur in blood clotting systems, visions systems,
and energy metabolism systems to name just a few.
After all, according to the theory of evolution, very large and apparently
impossible tasks are broken up into manageable parts. This is Dawkins's
main argument in his book, Climbing Mount Improbable. An impossible
statistical cliff that cannot be scaled in a single bound by natural selection
is scaled in small little evolutionary steps. Actual laboratory
experiments have been put forward to support such a concept.What is especially interesting is that these laboratory experiments
actually work!It has been
demonstrated in real time that the addition of unique components onto the end of
a metabolic cascade is in fact possible.
The
evolutionary biologist Kenneth Miller described one such experiment in his
recent book, Finding Darwin’s God.Miller quotes an experiment
done in 1982 by professor Barry
Hall. 2In this
experiment Hall deleted part of a metabolic pathway in an E.
colibacterium. Using the proper environmental pressures, the bacterium
"evolved" the missing piece of its cascade back again! 3
This
missing piece was actually the tip of a long cascade that breaks down sugar
molecules and extracts energy from them.Glucose
is the main sugar utilized by this cascading pathway. There are of course other
simple sugars, such as galactose and fructose etc., that can enter this pathway
as well .Each of these also
requires a unique enzyme or enzyme pathway to convert them to something that can
enter the cascade.Then, just like
adding more events to Foghorn’s cascade, more events can be added to the sugar
cascade.You see, there are
different kinds of sugars.Some of
these sugars are more complex than glucose, but can be broken down into glucose
and/or one of the other more simple sugars that are already part of the existing
cascade.Once this breakdown
occurs, a complex sugar molecule becomes just another part of an extended sugar
metabolism cascade. The problem is that unique enzymes are needed to break
down complicated sugar molecules.Some
sugars may even need more than one unique enzyme to break them down to a point
where they can enter the established cascade.
However, the benefits of obtaining these enzymes are great. If
such cascades of complex sugar breakdown can be established, any bacterium with
such capabilities would survive better than its peers in an environment that was
rich in a particular complex sugar.
The
ability to evolve such advantageous enzymes would certainly enhance the survival
of the species. In fact,the "evolved" bacteria in Hall's experiments quickly outgrew those that had not
yet evolved the needed enzyme.Of
course, this is only natural.It is
the law of survival commonly known as "the survival of the fittest".Hall went on to demonstrate the evolution of two and even three
additional steps added on to the original sugar cascade.
Although
I do agree with Behe when he says that cascades are indeed irreducibly complex,
I do not agree with his assertions that all such cascades are theoretically or
even practically impossible to evolve via the mindless processes of random
mutation and natural selection.Cascades in living systems are certainly as complicated and even vastly
more complicated than the one that whacked Foghorn Leghorn, but even such
complexity does not seem to be completely out of
reach in certain cases.
As
already described, the removal of one part of a cascade may not destroy its
ability to perform. The removal of an
enzyme that allows the utilization of complex sugars does not eliminate this
cascade's ability to continue to break down glucose or galactose or fructose.
Even the removal of the enzyme needed to break down glucose itself is not vital
to the function of the rest of the cascade. Even though glucose can no
longer be utilized, fructose still can be, along with several other types of
sugars. A cascade is therefore reducible without the loss of all
function, but it is still "irreducible" as far as the function that it
just lost is concerned. For example, a minimum number of parts are needed
that have a fairly specific internal structure in order for a bacterium to be
able to utilize the lactose sugar for energy. This minimum part
requirement creates a degree of irreducible complexity. Not just any
series of parts will do. Specific protein "parts" are needed.
In fact, Hall's experiment illustrates this specificity very well.
Hall
did delete the gene needed to produce an enzyme (lactase) that broke down the
sugar lactose into two other sugars called glucose and galactose. Both
glucose and galactose are part of the established sugar metabolism cascade of E.
coli. So obviously, without lactase, an E. coli bacterium can
no longer utilize lactose for energy. Hall deleted that lactase genes to
see if the E. coli bacteria would "evolve" back the ability to
utilize lactose when grown on a lactose enriched media. Hall's experiments were
a stunning and dramatic success. His colonies of E. coli quickly
"evolved" the ability to use lactose. There is just one little
catch.Hall did not
delete a spare tire gene (the evolved beta-galactosidase gene - ebg)
that required just one point mutation to produce an enzyme with a fairly high
level of lactase activity. But what if the
E. coli had not been so fortunate as to have this spare tire gene?
What would have happened then? Hall wondered about this himself. He
then deleted the spare tire gene as well as the original lacZ genes.
Would there be lactase evolution now? So far, none of these large
populations with high mutation rates has ever evolved the lactase ability
despite being subjected to lactose enriched selective media for tens of
thousands of generations. Evidently, no other gene could lend
its information by itself or in combination with any other gene to aid in the
bacterial "evolution" of lactase.Hall described these particular bacterial colonies as having "limited
evolutionary potential." 3But
why did these colonies have limited evolutionary potential?
It
turns out that there are statistical gaps that separate unique protein/enzymatic
functions from each other.Not
every protein sequence will be recognized as "beneficial" by a given
bacterium.In fact, the vast
majority of possible protein sequences required for the function of a particular
enzymatic activity, like the lactase function, will not be able to produce this
specified function - even a little bit.Because
of this problem, if proteins are not already very very close in sequencing to
begin with, the statistical odds that one will simply "evolve" into
another are remote because they are separated by a vast number of "neutral" amino acid sequences.Neutral
proteins cannot be guided by natural selection along any evolutionary path
whatsoever.Why?Because nature only sees functional differences.Nature cannot guide if it is blinded by functionally neutral differences.Thus, nature cannot guide evolution across neutral gaps.Obviously then, without this guidance of natural selection, evolution
simply stalls out after an endless "random walk" and dies.
The
results of Hall's experiment certainly prove Behe's point that at each step in a
cascade the required protein-enzyme is, in itself, irreducibly complex to at
least some degree. It also proves that this level of complexity is just
plain out of reach for many kinds of bacteria (not just the double mutant E.
coli studied by Hall).A
minimum number of amino acids are needed in a specific arrangement in order for
a specified function to be realized. If such an enzyme is altered or
reduced beyond a certain point, the specified function will simple vanish
completely. The genome that is left may not have what it takes to cross
the resulting neutral gap in function between what is and what is needed.
It is this neutral gap in function that forms the basis of Behe's argument that
irreducible complexity poses a significant hurdle for the theory of evolution.
Of
course, a cascade is no more complicated than the most complicated link in its
chain.If this most complicated
link can be overcome, then the rest of the chain would be easy to make.The question is, can the most complicated link be overcome in a
reasonable amount of time?
Professor Hall showed how simple bacteria can sometimes evolve links in a
cascade chain (but not always). If these
links were all insignificant hurdles they could simply be added up to
produce something quite significant - like a blood clotting cascade or a
metabolic pathway. But, as previously described, there might be a few
snags along the way if the links themselves are simply too irreducibly complex,
in and of themselves, to be overcome by a given life form in a given environment
- even in 4 billion years.
Professor
Hall never evolved anything that crossed a neutral gap that was more than two
mutations wide.The single
non-functional gap of two mutations that he did cross, he could not explain.In fact, by his own calculations, he figured this feat to be impossible -
taking an average of 100,000 years to cross.The apparent success of the crossing of even this tiny gap of
non-function astounded him.He
attempted to explain the success of this crossing by saying, "under
some conditions spontaneous mutations are not independent events." 3He went on to say that this is, "heresy, I am aware."If it is difficult for professor Hall to imagine the crossing of such a
small neutral gap, what would it take to cross a three mutation gap… or a four
mutation gap?
As
it turns out, Hall was mistaken in his calculations. He based his
estimates on a mistaken understanding of how the statistics of random walk
works. Hall assumed that each new mutation along the road toward the
desired beneficial mutation would have to become "fixed" or
transferred to the entire population before the next mutation could arrive in an
additive way. To understand how this need not be the case, consider the
following hypothetical situation:
Starting
with a steady state population of a trillion trillion (1024) random
sequences of 200 amino acids, each mutating every second into a new
sequence, how long would it take, on average, to find any one of the 10e200
lactose cleaving proteins? Surprisingly, it would take around 500 trillion
trillion years to find even one of the 10e200 lactase sequences on average.
In order to solve this problem,
one must first calculate how many different possible proteins could be made with
a series of 200 amino acids. The answer is on the order of 10e260 (1 with
261 zeros following). Compared to this number, even an astronomically
large number like 10e200 is relatively minute.What
happens is that each of the 10e200 functional proteins is surrounded by
trillions of non-functional protein sequences.
The
"potential space" of 10e260 options can be visualized as a gigantic
chess board. Each square on the chess board represents a different amino
acid sequence. Each member of a population can only occupy one square at a
time. A limited population simply cannot cover all the potential squares
on the chess board at any given moment of time. With each mutation to an
individual, it changes squares. If any one individual comes across a
beneficial sequence, like the lactase enzyme sequence, that individual and its
offspring will tend to stay on that square because of the selective advantage
given by that square. This advantage will be translated into an increased
population on and immediately around that particular square of the chess board.
The
interesting thing about random walk is that with each doubling of the length of
the average random walk, the time involved increases by a factor of 2. For
example, if the average random walk required for a particular colony of bacteria
to achieve a particular level of complexity required 5 neutral steps or changes
in DNA, the total number of options or potential space on our chess board would
be 4 to the power of 5 or 1,024 squares. Depending on our population's
size and mutation rate, we could estimate an average time required to reach all
of these squares at least once beginning at a random starting point. The
bigger our population, the faster we could reach all the squares.
For
instance, if we started out with a population of 1 trillion bacteria and if all
of these bacteria started out on one square on our chess board, it would take
around 65,000 generations for them to reach equilibrium over all the squares of
the chess board. At equilibrium, about 0.098% of them will be on each one
of the squares of the chess board. Even though 0.098% doesn't seem like a
big number, it actually works out to be 9.8 billion out of a population of 1
trillion. In other words, after 65,000 generations, there would be an
average of 9.8 billion bacteria covering each of the 1,024 squares on our chess
board of potential space. So obviously a gap of 5 neutral mutations would
not be a problem for a population of 1 trillion bacteria to cross in relatively
short order. But, what happens if we double the gap to 10? A gap of
10 mutations/steps would create a chess board with over 1 million squares of
potential space (1,048,576 to be exact). At equilibrium, our population of
1 trillion would have only 953,674 individuals on each of the squares instead of
the 98 billion it had when the gap averaged only 5 steps wide. Doubling
the gap again to 20 steps makes our chess board grow a million fold to just over
1 trillion squares of potential space (1,099,511,627,776). Now, our
population of 1 trillion would average a bit less than one member of the
population on any one square at any given point in time. I think the trend
is obvious by now, but just for kicks, doubling the gap again to 40 steps
increases the size of our chess board a trillion fold to just over 1 trillion
trillion squares. Now, each of the members of our population of one
trillion are surrounded, on average, by one trillion empty squares that they
have to search out all by themselves.
On
average then, each one of the members in our population will have to experience
over 1 trillion mutations (with none of them repeating) in order to reach all of
the potential spaces on our chess board. The time required for this
process truly reaches astronomical proportions in short order. With each
doubling of the neutral gap, the average time required increases by a factor of
two. The only way to reduce the required time is to increase the
population's size by a factor of two. This does help for a while, but very
quickly the required size of the population needed to keep up becomes
impractical for any environment to support and further evolution simply stalls
out.
Consider
now that many enzymes/proteins are hundreds or even thousands of amino acids in
length - and very specified in internal structure.The problem seems clear.In
fact, because of this problem all living things may have very "limited
evolutionary potential" indeed!Bacterial
colonies such asSalmonella,
Proteus, and Pseudomonas as
well as Halls double mutant E. coli colonies can be grown on Hall’s
selective media or any selective media in any sequence until the cows come home
and none of them will ever evolve the lactase enzyme. If this relatively
simple function does not evolve in certain creatures with "limited
evolutionary potential" what about functions that require multiple proteins
or systems working together simultaneously? How would such a multi-protein
function evolve gradually? For example, bacterial motility can come in
many different forms to include flagella, cilia, undulating membranes etc.
However, all known motility systems require many protein "parts"
working together simultaneously in a specified arrangement with each other (much
like the specified arrangement of amino acids required by single-protein enzymes
to work - only on a much larger scale of complexity). If it is often
difficult for various life forms to evolve single protein enzymes with
relatively few specified amino acids, imagine the difficulty required to evolve
a multi-protein function where entire proteins are specified in their
arrangement with each other. In fact, the odds are so bad that even on
paper the evolution of such a multi-protein function is simply beyond any
reasonable explanation - not to mention the fact that such a multi-protein
system of function has never been observed to evolve in real time in either the
laboratory or in nature.
For
example, an electric motor is irreducibly complex and it is not a cascading
system.It has a minimum number of
parts working together at the same time, in a specified orientation, in
order for it to have a function.It
needs a wire coil, a surrounding magnet, electrical current and bushings to
switch the electrical current back and forth at the proper moment in time.All these parts must of course be set up in the proper relation to each
other.However, if one part is
taken away, none of the other parts will "work" together - period.There is no "cascade" of function since all the parts work
together at the same time.So, in
order to get any function whatsoever from the electric motor, all the needed
parts must come together in a highly ordered/specified way - suddenly. An
electric motor, minus one of its parts, has no function. It might as well
be sitting in a junk pile at the city dump.
What
is especially interesting
is that some bacteria, as Behe points out, have little mechanical motors that
are very similar to electric motors.Just
like mechanical motors they require a minimal number of protein parts (each of
which are in turn made up of hundreds or even thousands of amino acid
"parts" in specified order), all working together at the same time, in
a specified orientation, to produce their rotary motion. If one adds up
all the required amino acids as "parts", literally hundreds of
thousands of parts are required in a specified order before function can be
realized.
Of
course, we are talking here about the famous bacterial flagellum
illustrated above.Flagella
are long whip-like cords attached to certain bacteria by a little motor of
sorts.1The motor
actually spins and causes the flagella to spin.The spinning flagella propels the bacterium through the liquid that it is
swimming in.The problem here is
that the motor that spins the flagella is not only "irreducibly
complex", but has a very high level of specified complexity (a large number
of required parts all working together at the same time).It is made up of a fair number of fully formed specified proteins (50 or
60 different ones), which are in turn made up of hundreds or thousands of
specified amino acids. Many of these parts are irreducible. If even
one of these irreducible parts is removed. The flagellar motor will not
work, not even a little bit.
Each
of these parts is, in itself, complex - just as each link in the cascade system
is complex.However, what separates
this type of irreducibly complex system from a cascading system is that each of
the parts is required to be in a specific orientation with all the other parts.
This adds another specified constraint to the system, which raises the level of
functional complexity significantly. In other words, a lot more
information is required to code for a flagellum than for a cascade of enzymes of
equal size and number. This creates an even larger neutral gap between
functions at this level of complexity as compared with single protein enzymes or
cascading systems. Again, no such multi-protein function has ever been
seen to evolve in real time. It just seems to be too far beyond what the
mindless processes of evolution are capable of - even with trillions and
zillions of years of time.
Some
do try and explain flagellar evolution by proposing that many of the parts in a
flagella are used as parts in other cellular systems of function. For
example, the actual flagella is very similar in structure to long tube-like
secretory structures. In fact, it is thought that some flagella might
function as both a motility structure as well as a secretory structure.
Likewise, many of the other parts in a flagellar motility system do other
similar jobs in other systems within the cell. Therefore, it seems obvious
to many that all these various parts already existed and therefore might easily
come together to form the motility system diagrammed above. The problem is
that the parts do not naturally self-assemble into a flagellar system, or any
other system of function. They must be directed in their assembly by a
pre-formed coded message contained in that cell's DNA. It is like the
parts of a car. Many of the same parts could be used to build a boat, or a
house, or any number of other things. However, planning is needed because
the parts themselves, if left to themselves, do not self-assemble to form any
one of these things (even if given plenty of energy and opportunity to interact
with each other). Likewise, starting with a bacterium without a motility
system but with all the needed parts to make one, there is no series of point
mutations that will get it from what it has to the goal of motility without the
crossing of neutral gaps in function (despite the selection pressure or
advantages of motility should this function happen to evolve). In other
words, there is no way to mutate the genetic code were each and every mutation
will be beneficially advantageous until motility is achieved.
So,
Miller’s attempt to dismiss Behe’s challenge of irreducible complexity
stands on very shaky ground. At best it is limited to the level of single
protein enzymes and relatively simple cascading systems of single protein
enzymes.Even these examples fall very short as cascading systems are far less
complex than multi-protein systems where all the protein parts work together at
the same time in a specified arrangement.
In many cases, even these lower level cascading functions pose significant
hurdles that severely limit the mindless processes of naturalistic evolution.
Obviously then, it is only intuitive that anything of even slightly greater
complexity might just stall out the processes of evolution completely.
And, in real life, this is exactly what we observe. Again, as far as I am
aware, no life form in any environment or series of environments has ever been
shown to evolve any multi-protein system of function where the proteins all work
together at the same time in a specific orientation with each other.
So,
even though some irreducible functions can evolve and have evolved, there is a
ladder of irreducible complexity that quickly moves beyond what evolution has
done and can do. In my opinion then, Behe's concept of irreducible
complexity remains pretty much untouched as a challenge to the modern theory of
evolution and as a strong voice in favor of intelligent design.
Behe,
Michael J. Darwin's Black Box,
The Free Press, 1996.
Miller,
Kenneth R., Finding Darwin's God,
HarperCollins Publishers, 1999.
B.G.
Hall, Evolution on a Petri Dish.The
Evolved B-Galactosidase System as a Model for Studying Acquisitive Evolution
in the Laboratory, Evolutionary Biology, 15(1982): 85-150.
 Well,
we must first ask if Rube Goldberg machines really are "irreducibly
complex"? It is true that if one
part is removed the cascade is interrupted at that point. It is kind of
like taking a domino out of a line of dominos that are balanced on their ends.
The dominos after this point will not be knocked over by the previously
cascading dominos. In this way, the final
function of the cascade is rendered useless. However, the cascade is in
fact still able to function from the point of interference onwards. So,
its function cannot be said to be completely destroyed just because of a single
point of interruption. Also, by the very nature of cascades, more
parts can be added on the originating end of the cascade to make it longer and
longer still... like adding more and more dominos onto the end of a chain of
dominos. At first one starts with one domino, then two, three, four and so
on until one has a very long cascade set up.
Well,
we must first ask if Rube Goldberg machines really are "irreducibly
complex"? It is true that if one
part is removed the cascade is interrupted at that point. It is kind of
like taking a domino out of a line of dominos that are balanced on their ends.
The dominos after this point will not be knocked over by the previously
cascading dominos. In this way, the final
function of the cascade is rendered useless. However, the cascade is in
fact still able to function from the point of interference onwards. So,
its function cannot be said to be completely destroyed just because of a single
point of interruption. Also, by the very nature of cascades, more
parts can be added on the originating end of the cascade to make it longer and
longer still... like adding more and more dominos onto the end of a chain of
dominos. At first one starts with one domino, then two, three, four and so
on until one has a very long cascade set up.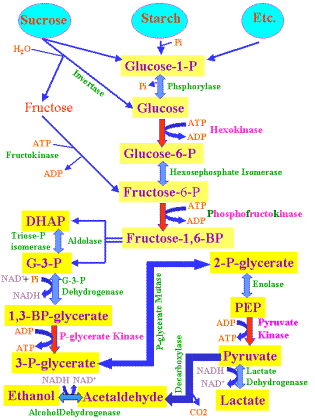 The
evolutionary biologist Kenneth Miller described one such experiment in his
recent book, Finding Darwin’s God.
The
evolutionary biologist Kenneth Miller described one such experiment in his
recent book, Finding Darwin’s God.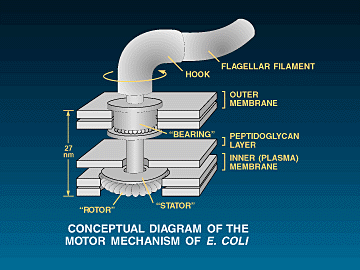 What
is especially interesting
What
is especially interesting