Natural
Selection
Sean
D. Pitman M.D.
©
July 2003
 According to evolutionary theory, natural selection is the driving force
behind the process of evolution.
The mindless processes of nature are thought by most modern scientists to be the
creative forces the mold and govern everything that we see in and around us - to
include all life forms. Truly then, Nature is the Creator. Nature is
a mindless god of sorts. But, by what power does this god create all the
fantastic variety and creativity that we see around us? - especially when we
look at the workings of living things?
According to evolutionary theory, natural selection is the driving force
behind the process of evolution.
The mindless processes of nature are thought by most modern scientists to be the
creative forces the mold and govern everything that we see in and around us - to
include all life forms. Truly then, Nature is the Creator. Nature is
a mindless god of sorts. But, by what power does this god create all the
fantastic variety and creativity that we see around us? - especially when we
look at the workings of living things?
As far as the variety of life forms are concerned, the theory of evolution
proposes that the nature created and is still creating the huge variety
of living creatures via random genetic mutations that can be selected as
advantageous or disadvantageous in particular competitive environments over vast
spans of time.
Nature is said to do this by preferentially selecting those life forms
that survive and pass on their genes to their offspring the best.
This "natural selection" uses competition for survival.
The production of slightly different offspring provides the opportunity
of sorting out advantageous traits from the less advantageous ones.
More and more traits are added or subtracted in this way until a
creature's offspring are fine tuned to the particular environmental niche that
they occupy. The whole process is
often referred to as, "The Survival of the Fittest."
This is rather a neat idea, but
exactly how does it work? Does natural
selection tend toward genetic diversity or stability?
 A
famous example of natural selection is England's peppered moth (Biston
betularia). Despite the current debate on the actual methods of the original
study it is still a good example to illustrate natural selection.
In the original study by H.B. Kettlewell, he was wondering why the
peppered moth was more commonly dark than specimens that had been collected and
saved from earlier times in England’s history.
The peppered moth had been generally much whiter in color, but now it was
much blacker. Why this change?
It was suggested that when the industrial revolution arrived in England,
the pollution had turned the bark of the trees a much darker color.
Since light colored moths are much easier for birds to see on a dark
background, they were preferentially eaten.
However, the few darker moths survived better to pass on their darker
coloring to their offspring. Pretty
soon, there were lots of dark moths and very few
light colored moths - or so the story went. In reality, England's peppered
moths do not generally rest on tree trunks (despite what some of the staged
pictures produced by Kettlewell seem to suggest). However, subsequent
studies seemed to confirm Kettlewell's main hypothesis that England's peppered
moths were in fact getting darker over time. Still, this is
still not necessarily an example of evolution in action.
A
famous example of natural selection is England's peppered moth (Biston
betularia). Despite the current debate on the actual methods of the original
study it is still a good example to illustrate natural selection.
In the original study by H.B. Kettlewell, he was wondering why the
peppered moth was more commonly dark than specimens that had been collected and
saved from earlier times in England’s history.
The peppered moth had been generally much whiter in color, but now it was
much blacker. Why this change?
It was suggested that when the industrial revolution arrived in England,
the pollution had turned the bark of the trees a much darker color.
Since light colored moths are much easier for birds to see on a dark
background, they were preferentially eaten.
However, the few darker moths survived better to pass on their darker
coloring to their offspring. Pretty
soon, there were lots of dark moths and very few
light colored moths - or so the story went. In reality, England's peppered
moths do not generally rest on tree trunks (despite what some of the staged
pictures produced by Kettlewell seem to suggest). However, subsequent
studies seemed to confirm Kettlewell's main hypothesis that England's peppered
moths were in fact getting darker over time. Still, this is
still not necessarily an example of evolution in action.
 Gregor
Johann Mendel (1822-1884), an Austrian monk and a contemporary of Darwin, is
generally recognized as the father of modern genetics and the study of genetic
inheritance. Mendel showed
conclusively that the genes (alleles) of creatures contained built-in abilities
for inherited variation that are not dependent upon mutational changes
but upon the built-in variety potential of a given gene pool of allelic options.
He found that
some
allelic traits were "dominant" while others were "recessive."
Each trait was coded for by two separate allelic codes on equivalent
positions on two separate but matching chromosomes.
As long as one of the codes was the dominant code, it would block out the
other code. So, an individual with
two different codes for the same allelic positions would express the dominant
trait. For example,
if one parent had two dominant color codes and the other parent had two
recessive color codes at the allelic position coding for hair color, then the
offspring would all be the dominant color (i.e., brown vs. blond).
However, if each parent had one dominant and one recessive color code
then, according to the calculated odds of inheritance, three-quarters of the
offspring would be dominant in color and one-quarter recessive (The offspring
with both dominant and recessive codes would express the dominant color.
Only those offspring with both recessive codes would express the
recessive color). If both parents
had both recessive color codes then all the offspring would have the recessive
color.
Gregor
Johann Mendel (1822-1884), an Austrian monk and a contemporary of Darwin, is
generally recognized as the father of modern genetics and the study of genetic
inheritance. Mendel showed
conclusively that the genes (alleles) of creatures contained built-in abilities
for inherited variation that are not dependent upon mutational changes
but upon the built-in variety potential of a given gene pool of allelic options.
He found that
some
allelic traits were "dominant" while others were "recessive."
Each trait was coded for by two separate allelic codes on equivalent
positions on two separate but matching chromosomes.
As long as one of the codes was the dominant code, it would block out the
other code. So, an individual with
two different codes for the same allelic positions would express the dominant
trait. For example,
if one parent had two dominant color codes and the other parent had two
recessive color codes at the allelic position coding for hair color, then the
offspring would all be the dominant color (i.e., brown vs. blond).
However, if each parent had one dominant and one recessive color code
then, according to the calculated odds of inheritance, three-quarters of the
offspring would be dominant in color and one-quarter recessive (The offspring
with both dominant and recessive codes would express the dominant color.
Only those offspring with both recessive codes would express the
recessive color). If both parents
had both recessive color codes then all the offspring would have the recessive
color.
So
you see, such a method of option swapping at set locations on chromosomes allows
for a huge variety of expressed morphologies or phenotypes.
However, since the codes only swap with each other at set locations, the body
parts themselves maintain their usual orientation with each other. In
other words, genetic
recombination will not cause an eye to grown on a baby's foot or an ear to grow
inside its stomach. The swapping of options is random, but limited as to
exactly where the swapping can and cannot take place. For example, it
works much like interchangeable parts on a car. If I don't like the
hubcaps on my car, I can swap them out for new ones that still fit in the same
location, but they may look very different. I can also swap out the
steering wheel for a very different looking one, but it still fits in exactly
the same place on the car and it does pretty much the same job.
This
same thing happens during genetic recombination. One particular position
on a chromosome might code for eye color. Many different interchangeable
parts or codes for eye color have the potential to occupy and "work"
in this spot. But, codes for eye color would not work so well if they were
placed where the code for nose size is supposed to go. This would be like
trying to put a hubcap where the carburetor is supposed to go. Some parts
are simply not interchangeable, and the process of genetic recombination knows
this. So,
during the process of genetic recombination, the genes or allelic options
themselves have not been changed, just their expression (i.e., The hubcaps,
steering wheels, carburetors, and all other part options are still the
same. It is just that some parts are used to make some car
"expressions" while other part types are used to make other car
"expressions").
There
are many different variations and complications, but this is the basic idea that
Mendel discovered. In other words,
great diversity can be had through a selection process that picks between
previously established traits or allelic part
options. This is why breeding has been such a thriving occupation
for centuries. Selective breeding based on the potential of
genetic recombination alone (for the most part) is responsible for the great
varieties of cats, dogs, horses, and even humans, as well as a host of other
breeds - to include Darwin’s famous finches.
Mendelian genetics and other types of genetic recombination are the
primary reason why children do not look exactly like their parents or each other
(unless they happen to be twins). 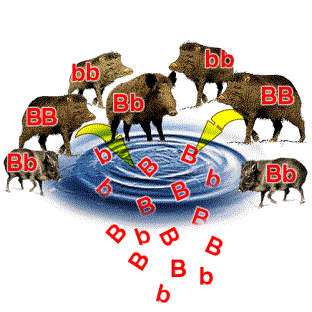 Children
from the same set of parents do have variations in their
appearance and yet these variations are dependent upon nothing more than different
expressions and mixes of the same identical gene options from both parents (the
parental gene pool of options). Note
carefully thought that this process (as it ideally works) has nothing to
do with mutation so nothing new as far as the "gene pool potential" is
concerned is created. No unique genetic functions are evolved during this
process (i.e., An exchange of one type of hubcap for another type of hubcap that
already existed in the pool of hubcap options does not make the pool of options
change).
Children
from the same set of parents do have variations in their
appearance and yet these variations are dependent upon nothing more than different
expressions and mixes of the same identical gene options from both parents (the
parental gene pool of options). Note
carefully thought that this process (as it ideally works) has nothing to
do with mutation so nothing new as far as the "gene pool potential" is
concerned is created. No unique genetic functions are evolved during this
process (i.e., An exchange of one type of hubcap for another type of hubcap that
already existed in the pool of hubcap options does not make the pool of options
change).
A
mutation, on the other hand, is a change in a specific gene that was not
originally inherited. The different
white and black colors in the peppered moth are clearly the result of genetic
recombination at work. The peppered
moth gene pool already contained codes for both the light and the dark colors of
peppered moths. Nature did in fact
play a role in selecting the most common color from this pre-established pool of
options but it did not create the options in this case (at least not in
observable time). So, such examples of changes do to genetic recombination
cannot be used as examples of evolution in action because clearly, they are not
making anything new as far as the gene pool is concerned.
For example, no matter how much selection pressure is applied to England's
peppered moth, a green peppered moth will never evolve via genetic recombination
alone. Why? Because the code for green color is not in the peppered
moth's genetic pool of options (i.e., If there are no green hubcaps at Wal-Mart,
where I do my hubcap shopping, my car will simply have to do without green
hubcaps). The only way to get this option to come into the gene pool of
options is for random mutations or mistakes in the genetic code to create this
option de novo.
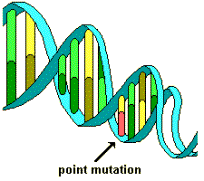 The
problem is that random mutations generally limit existing genetic functions and
so nature almost always selects against the changes produced by random
mutations. In other words, if an
individual sustains a mutation in one of its genes, this mutation lessens the
function of this gene most of the time. With
a less functional gene, the creature has a higher probability of competing less
well than peers. This mutant
creature and its offspring will most likely die off because of this deficiency.
Of course, after the die off occurs, this particular mutation would be removed
from the gene pool of options for this "kind" of creature.
In this way, nature in fact limits mutational change to the genes of most
creatures. For many
creatures, such as mammals, mutations are fairly rare to begin with; on the
order of one mutation per gene per 100,000 generations. 1
Understand also that the majority of even this relatively small
number of mutations are detrimental in nature (The ratio of detrimental vs.
beneficial mutations is thought to be around 1000:1 - with the rest being
"neutral" as far as function is concerned). One major problem is
one of how to eliminate the detrimental mutations as fast or faster than the
much lower rate of beneficial mutations. For humans this is turning out to
be quite a mysterious problem. In fact, it seems that the human species
may be deteriorating at an alarming rate. Consider
an excerpt from a fairly recent Scientific American article entitled,
"The Degeneration of Man" :
The
problem is that random mutations generally limit existing genetic functions and
so nature almost always selects against the changes produced by random
mutations. In other words, if an
individual sustains a mutation in one of its genes, this mutation lessens the
function of this gene most of the time. With
a less functional gene, the creature has a higher probability of competing less
well than peers. This mutant
creature and its offspring will most likely die off because of this deficiency.
Of course, after the die off occurs, this particular mutation would be removed
from the gene pool of options for this "kind" of creature.
In this way, nature in fact limits mutational change to the genes of most
creatures. For many
creatures, such as mammals, mutations are fairly rare to begin with; on the
order of one mutation per gene per 100,000 generations. 1
Understand also that the majority of even this relatively small
number of mutations are detrimental in nature (The ratio of detrimental vs.
beneficial mutations is thought to be around 1000:1 - with the rest being
"neutral" as far as function is concerned). One major problem is
one of how to eliminate the detrimental mutations as fast or faster than the
much lower rate of beneficial mutations. For humans this is turning out to
be quite a mysterious problem. In fact, it seems that the human species
may be deteriorating at an alarming rate. Consider
an excerpt from a fairly recent Scientific American article entitled,
"The Degeneration of Man" :
According
to standard population genetics theory, the figure of three harmful mutations
per person per generation implies that three people would have to die
prematurely in each generation (or fail to reproduce) for each person who
reproduced in order to eliminate the now absent deleterious mutations [75% death
rate]. Humans do not reproduce fast enough to support such a huge death
toll. As James F. Crow of the University of Wisconsin asked rhetorically,
in a commentary in 'Nature' on Eyre-Walker and Keightley's analysis: "Why
aren't we extinct?"
4
Such
problems certainly are a challenge for natural selection to keep up with and get
rid of much less overcome to use in some fantastically beneficial way.
Clearly natural
selection is a real force of nature, but it is limited by the genetic options
that it can pick from. New options
can only be added through mutation of the original options - and this seems to
be very limited at best.
 But
what about some famous mutational benefits that have survived?
What about sickle cell anemia for example?
Sickle cell anemia is caused by a single "point" mutation of
the hemoglobin molecule. This
molecule is responsible for oxygen transport in red blood cells.
When it has the mutation that causes sickle cell anemia, it does not
carry oxygen as well. It also has a
tendency to "sickle" or polymerize into a shape that is inflexible and
unable to pass through very small vessels called capillaries.2
This causes the individual with sickle cell disease a great deal of
pain and eventually an early death. So,
why has nature preserved or "selected" to maintain this mutation in
our particular human gene pool of options?
Nature has done this because of another even more deadly problem called
malaria. Malaria is caused
by a microscopic protozoan that lives part of its life cycle as a parasite
in human
But
what about some famous mutational benefits that have survived?
What about sickle cell anemia for example?
Sickle cell anemia is caused by a single "point" mutation of
the hemoglobin molecule. This
molecule is responsible for oxygen transport in red blood cells.
When it has the mutation that causes sickle cell anemia, it does not
carry oxygen as well. It also has a
tendency to "sickle" or polymerize into a shape that is inflexible and
unable to pass through very small vessels called capillaries.2
This causes the individual with sickle cell disease a great deal of
pain and eventually an early death. So,
why has nature preserved or "selected" to maintain this mutation in
our particular human gene pool of options?
Nature has done this because of another even more deadly problem called
malaria. Malaria is caused
by a microscopic protozoan that lives part of its life cycle as a parasite
in human 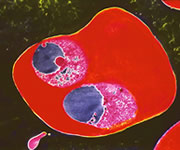 red blood cells. However,
the malaria parasite needs a specific oxygen concentration to survive.
Those humans with sickle cell anemia cannot maintain the necessary oxygen
level needed for the malaria parasite to survive.
So, those with sickle cell anemia, or even sickle cell "trait"
(one sickle cell allele - instead of the two possible sickle cell gene alleles)
survive better in an environment where there is malaria.
However, this survival comes at a high cost.
They have a less functional hemoglobin molecule.
A loss of function or a disruption of previous functions or interactions is far
easier to achieve via random mutations than a new function that is not dependent
upon the loss of or interference with any pre-established functions or
interactions.
red blood cells. However,
the malaria parasite needs a specific oxygen concentration to survive.
Those humans with sickle cell anemia cannot maintain the necessary oxygen
level needed for the malaria parasite to survive.
So, those with sickle cell anemia, or even sickle cell "trait"
(one sickle cell allele - instead of the two possible sickle cell gene alleles)
survive better in an environment where there is malaria.
However, this survival comes at a high cost.
They have a less functional hemoglobin molecule.
A loss of function or a disruption of previous functions or interactions is far
easier to achieve via random mutations than a new function that is not dependent
upon the loss of or interference with any pre-established functions or
interactions.
The
evolution of new genetic functions that relies on the disruption of
pre-established functions is a quick and easy process that happens all the
time. The evolution of antibiotic resistance to all the various
antibiotics that we have developed is based on this process. The de
novo development of the antibiotic function in a colony of bacteria is dependent
upon the fact that all antibiotics are quite specific in their interactions with
particular intra-bacterial target sequences. Any little disruption of the
target sequence will interfere to one degree or another with the antibiotic's
interaction with this sequence. This interference results in
equivalent resistance to the antibiotic. Obviously then, the ratio of
interfering mutations involving this target sequence is
very large when compared to the total number of potential mutations that could
affect this sequence. This high ratio of what will "work"
compared with what will not "work" creates a very small gap between
what is and what might be more helpful in the current situation or
"environment".
The
problem is that there are other cellular functions that are not dependent upon
the disruption of a pre-established functions or interactions. For
example, single protein enzymes act alone, without the need for the loss or
interference of any other cellular function. Several such enzymes have
been shown to evolve de novo in living organisms such as bacteria.
However, given the much higher specificity of the average enzyme (as compared to
antibiotic resistant target sequences), it is much harder to evolve a new
beneficial enzyme than it is to evolve the much simpler antibiotic-type
function. Some examples of enzyme evolution include the evolution of the lactase
and nylonase functions in bacteria.3
However, such examples are extremely limited and highly constrained by both
environment as well as the starting genetic real estate of the population of
bacteria. Many if not most types of bacteria simply cannot evolve certain
specific enzymatic function regardless of the size of their population, high
mutation rates, and tens, hundreds, or even millions of generations of
time. Of course, the problems only get worse from here on out.
Functions of far greater complexity exist inside living cells. For
example, many functions require the simultaneous action of multiple proteins
interacting in a specified arrangement with each other. If it is difficult
to evolve just one relatively simple enzymatic-type function, imagine how hard
it would be to evolve any function of the level requiring such a specific
multi-protein system as is used in various bacterial systems of motility (i.e.,
the flagellum - requires 50 or 60 unique proteins all working together at the
same time in a specific orientation with each other). Interestingly
enough, no such function that requires multiple proteins working in specified
concert has ever been shown to evolve in real time. It just doesn't happen
even when all the required parts needed to produce some fabulously beneficial
function are already present inside the same cell as parts of other systems of
function. The problem is that the parts themselves, if left to themselves,
just don't know how to self-assemble to form much of anything. They need
outside information telling them how to assemble in specified ways to produce
higher and still higher levels of functional complexity. Without this
outside information, in the form of pre-established genetic codes or the insight
of an intelligent mind, the parts are no more creative in their interactions
than a junk pile during an earthquake (even if that earthquake lasts for
millions, billions or even trillions of years - - nothing more complex
than a pile of random parts will be realized).
So,
natural selection is a real force of nature.
However natural selection is very much limited to a pre-established gene
pool of options. Natural selection
generally interacts with Mendelian laws and other laws of genetic recombination
to create diversity. Mutations are
almost always detrimental and are therefore selected against, as a rule, by
natural selection. And, even the mutations
that are beneficial are limited to the lowest levels of functional
complexity. There simply are no examples of real evolution producing
anything of the level of complex function that requires multiple
proteins working in a specified orientation at the same time.
Natural
selection is therefore a force that generally acts against
the other natural force of evolution (random mutations) and even at its best is
not much of a help to the popular theory of evolution.
1.
Ayala, Francisco J. Teleological
Explanations in Evolutionary Biology, Philosophy of Science, March, 1970, p.
3.
2.
Stryer, Lubert. Biochemistry,
3rd ed., 1988, pp. 153, 744.
3.
B.G.
Hall, Evolution on a Petri Dish. The
Evolved B-Galactosidase System as a Model for Studying Acquisitive Evolution in
the Laboratory,
Evolutionary
Biology, 15(1982): 85-150.
4.
Beardsley, Tim, The Degeneration of Man,
Scientific American, April, 1999, p32
.
Home Page
. Truth,
the Scientific Method, and Evolution
.
Methinks
it is Like a Weasel
. The
Cat and the Hat - The Evolution of Code
.
Maquiziliducks
- The Language of Evolution
. Defining
Evolution
.
The
God of the Gaps
. Rube
Goldberg Machines
.
Evolving
the Irreducible
. Gregor
Mendel
.
Natural
Selection
. Computer
Evolution
.
The
Chicken or the Egg
. Antibiotic
Resistance
.
The
Immune System
. Pseudogenes
.
Genetic
Phylogeny
. Fossils
and DNA
.
DNA
Mutation Rates
. Donkeys,
Horses, Mules and Evolution
.
The
Fossil Record
. The
Geologic Column
.
Early Man
. The
Human Eye
.
Carbon
14 and Tree Ring Dating
. Radiometric
Dating
.
Amino Acid
Racemization Dating
. The
Steppingstone Problem
.
Quotes
from Scientists
. Ancient
Ice
.
Meaningful
Information
. The
Flagellum
.
Harlen Bretz
Since
June 1, 2002
Debates:
Stacking
the Deck
God
of the Gaps
The
Density of Beneficial Functions
All
Functions are Irreducibly Complex
Ladder
of Complexity
Chaos
and Complexity
Confusing
Chaos with Complexity
Evolving
Bacteria
Irreducible
Complexity
Scientific
Theory of Intelligent Design
A
Circle Within a Circle
Crop
Circles
Mindless
vs. Mindful
Single
Protein Functions
BCR/ABL
Chimeric Protein
Function
Flexibility
The
Limits of Functional Flexibility
Functions
based on Deregulation
Neandertal
DNA
Human/Chimp
phylogenies
Geology
The
Geologic Column
Fish
Fossils
Matters
of Faith
Evidence
of Things Unseen
The
Two Impossible Options
Links
to Design, Creation, and Evolution Websites
Since
June 1, 2002
