
The Fossil Record
Sean D. Pitman M.D.
© May 2001
Updated October 2015
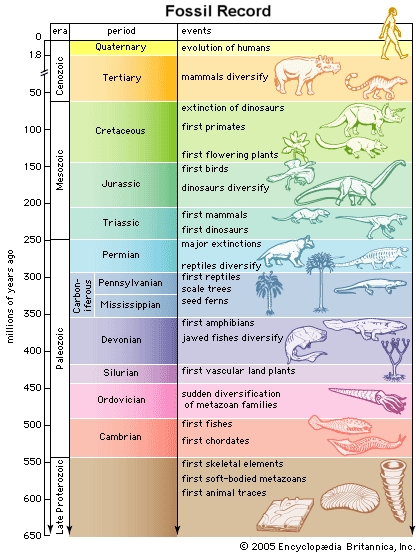
The standard approach to looking at fossils in the geological column is to assume that lower is older. Since the geologic column represents millions of years of Earth's history, then obviously the fossils in each of the layers must be the same age as the layer in which they are found. What is especially interesting is that the fossils do appear to show a progression from the most "simple" of organisms, such as single celled creatures like bacteria, to the most "complex" organisms, such as vertebrates, mammals, and of course humans. This evolutionary progression seems to be clearly demonstrated in that certain kinds of creatures in the upper layers are rarely if ever seen in lower layers. Many of the layers also show a certain specialization. Some layers contain mostly fish fossils while others contain land-dwelling creatures such as dinosaurs. Since each of these layers seems so specialized it is easy to conclude that one type of creature gave rise to the next type of creature over the course of whatever time it took to form the various layers between them. Radiometric dating and many other techniques are used to support the idea that this transformation process took tens and hundreds of millions of years.
For most scientists all of this seems so obvious that it is difficult to question. It goes against human nature to challenge long held ideas of truth. However, science is suppose to do just that - challenge ideas. Why? Because often what seems obvious initially does not turn out to be true. The scientific method is all about testing and retesting theories since no theory is ever proven by science. The scientific method can only disprove theories or increase the power of previous predictions that have yet to be disproved, but it can never absolutely prove anything to be absolutely true. Therefore, the best thing to do in science is to continually question and test previous hypothesis and theories to see if they continue to hold up under scrutiny. So, let's take another look at the available evidence and see if any other possibilities present themselves.
Surprisingly there are quite a few problems with the geologic column itself being a representation of millions of years of Earth's history. Much of the evidence available seems to point more toward the rapid formation of much of the column. Of course popular science disagrees stating that these layers represent millions of years of history and that the fossils they contain are likewise millions of years old. However, there are numerous features of both the fossil record and the geologic column that suggest another interpretation.
|
|
 What about the fact that the "simple" organisms are buried in
the lower levels and the more "complicated" ones are buried in the higher
levels? Doesn't this fact support the notion that simple
organisms evolved into more and more complex organisms over time, with the more
complex organisms buried and fossilized above the earlier and simpler life
forms? Certainly this seems like a very logical assumption. But, things
just aren't that easy. There are a number of potential problems with this
interpretation of the fossil record.
What about the fact that the "simple" organisms are buried in
the lower levels and the more "complicated" ones are buried in the higher
levels? Doesn't this fact support the notion that simple
organisms evolved into more and more complex organisms over time, with the more
complex organisms buried and fossilized above the earlier and simpler life
forms? Certainly this seems like a very logical assumption. But, things
just aren't that easy. There are a number of potential problems with this
interpretation of the fossil record.
For example, it is interesting to note that some general kinds of fossilized creatures are very generally found in the same relative vertical orientation, with respect to each other in the fossil record, that they would have naturally been found in during life. Single celled organisms make their first appearance in the lowest layers followed by multicelled ocean bottom-dwelling creatures like sponges and worms etc. Higher up come creatures like bony fishes, then land plants and animals, then birds and larger land animals.
Of course, this is a very general pattern and does not explain why certain creatures that lived on the bottoms of oceans, like trilobites, make their first appearance in the Cambrian (505-540 Ma) while other creatures that live on ocean bottoms, like crabs and lobsters, don't appear until the beginning of the Cretaceous (65-145 Ma).83 Why would creatures that would seem to share the same general environment while alive be so widely separated in the fossil record if they did indeed live at the same time and in pretty much the same location? If the geologic column truly represents a series of closely spaced catastrophic burial events instead of long ages of time, how can this feature be explained? Certainly this seems like a difficult and rather mysterious problem for those, like myself, who might think to question the long age notion of the fossil record.
At least a partial explanation might be found in the fairly recently discovered fact that at least some nested hierarchical patterns to the distribution of different populations (both living and within the fossil record) seem to be strongly related to ecological and population-size factors.
"The common pattern of species identities associated with species area relationships is the 'nested subsets' pattern. This pattern arises when species that appear on few islands occur only on the islands with the most species, while only the most widespread species are found on the islands with few species (Wright et al., 1998). The nested subset pattern arises because species differ in their distributions across space. Some species use a wider range of resources or persist across a wider range of habitats than others (Brown, 1984; Brown et al., 1996; Hanski and Gyllenberg, 1997). Generally, species that use a wide range of resources or tolerate a variety of abiotic conditions can establish more populations in more places than comparable species with relatively narrow niches (Brown, 1995; Cook and Quinn, 1995). Differences in the ability of species to distribute themselves across space have distinct consequences for the structure of communities. Sites that encompass a greater area tend to have more species (Rosenzweig, 1995). This is because large areas include a subset of species not found elsewhere. Therefore, the nested subset pattern of species distribution in space is thought to reflect the gradient in abundance among species (Gaston, 1996; Leitner and Rosenzweig, 1997; Maurer, 1999). . . [These features are consistent with the hypothesis of] "isolated habitat 'islands'." 109
Using this line of reasoning, one might reasonable hypothesize that trilobites appear in the fossil record before crabs and lobsters at least party because of the relative abundance of trilobites compared to crabs and lobsters. This hypothesis is at least plausible given the author's conclusion that, "Species identities and their relative abundances are non-random properties of communities that persist over long periods of ecological time and across geographic space. This is consistent with species abundance contributing heavily to evolutionary patterns." 108 After all, "It's very rare to find fossils of lobsters" ( Link ). General mobility, ability to survive catastrophic conditions, and other ecological/habitat factors could also reasonably contribute to the differential location of trilobites vs. lobsters and crabs in the fossil record. For example, coelacanth fish exist in the fossil record for what are thought to have been 400 million years. Then they suddenly disappear from the fossil record some 80 million years ago only to reappear alive an well swimming around in oceans today. Clearly, some types of coelacanths lived in habitats that did not lend themselves to fossilization while others did. Some habitats are clearly more susceptible to the preservation of fossils. If those specific habitats are not occupied by a particular kind of creature, it may not be preserved in the fossil record even though it is still alive and well in some other habitat. Consider also that the crayfish was once thought to have evolved from lobster-like ancestors around 140 Ma. This was until very modern-looking crayfish were subsequently found in sedimentary rocks dating up to 300 Ma ( Link ).
 This
brings up the interesting problem of the "first appearance" of all kinds of
fossils being pushed farther and farther back in supposed geologic time - often
dramatically so. For example, in January of 2008, Rudkin and his
colleagues, including Graham Young of the Manitoba Museum, spotted fossils of
horseshoe crabs buried in rocks thought to be 445-million-years-old from the
Ordovician period in central and northern Manitoba. They describe the discovery
in the January issue of the journal Paleontology. Of course, horseshoe
crabs are not true crabs. They more closely resemble spiders and scorpions
in their body plan. However, it is interesting that these creatures were thought
to exist no farther back than 350 million years (Carboniferous period) before
this discovery pushed them back another 100 Ma to 445 Ma. Both the Carboniferous
and the Jurassic fossil discoveries indicate the "ancient" horseshoe crabs
greatly resembled their modern-day counterparts. And, amazingly enough, analysis
of the recent finds also indicates the ocean creatures haven't changed much over
the eons. "We wouldn't necessarily have expected horseshoe crabs to look very
much like the modern ones, but that's exactly what they look like," Rudkin said.
"This body plan that they've invented, they've stayed with it for almost a half
a billion years. It's a good plan," Rudkin told LiveScience. "They've
survived almost unchanged up until the present day." (
Link
)
This
brings up the interesting problem of the "first appearance" of all kinds of
fossils being pushed farther and farther back in supposed geologic time - often
dramatically so. For example, in January of 2008, Rudkin and his
colleagues, including Graham Young of the Manitoba Museum, spotted fossils of
horseshoe crabs buried in rocks thought to be 445-million-years-old from the
Ordovician period in central and northern Manitoba. They describe the discovery
in the January issue of the journal Paleontology. Of course, horseshoe
crabs are not true crabs. They more closely resemble spiders and scorpions
in their body plan. However, it is interesting that these creatures were thought
to exist no farther back than 350 million years (Carboniferous period) before
this discovery pushed them back another 100 Ma to 445 Ma. Both the Carboniferous
and the Jurassic fossil discoveries indicate the "ancient" horseshoe crabs
greatly resembled their modern-day counterparts. And, amazingly enough, analysis
of the recent finds also indicates the ocean creatures haven't changed much over
the eons. "We wouldn't necessarily have expected horseshoe crabs to look very
much like the modern ones, but that's exactly what they look like," Rudkin said.
"This body plan that they've invented, they've stayed with it for almost a half
a billion years. It's a good plan," Rudkin told LiveScience. "They've
survived almost unchanged up until the present day." (
Link
)
This whole thing is turning into somewhat of a routine. Older and older specimens of various creatures are being found quite commonly and almost always their modern-day counterparts, if they exist, are essentially no different. How odd from an evolutionary perspective to find example after example of morphologic stasis over hundreds of millions of years of evolutionary time . . .
Other examples of fossils being dating much older than previously thought include:
Australopithecus
(ape thought to be ancestral to humans) dated 4 million years older than
previously thought (
Link
).
Platypus dating 40 million years older than previously thought (to 120 Ma)
pushing the first appearance of mammals back to the time of the dinosaurs (
Link
).
A block of amber (fossil resin) encasing an extinct, stingless bee (Proplebeia
dominicana) carrying a clump of orchid pollen on its back shows that
these "masterpieces" among flowers appeared sometime between 76 million and
84 million years ago, much earlier than previously believed. (
Link
).
Orb-weaving spiders arrived during the Jurassic period, not the Cretaceous
period, making them about 100 million years older than has been believed,
the Times of London reported Wednesday (
Link
).
Ants 40 million years older than previously thought (
Link
).
Animal fossils associated with the Ediacaran Period (635 to 543 million
years ago) have been found in sediments that date squarely in the
Proterozoic Eon, 1.6 billion years ago - a billion years older than
previously thought (
Link
).
The phylogenetic group of crustaceans includes nearly 100,000 species,
including relatively big creatures such as prawns, lobsters and crabs, but
also smaller specimens such as sea fleas and bay barnacles. In geological
terms, the group is as much as 480 million years old; some rare fossils even
belong to the late Cambrian and date as far back as 505 million years ago.
The new fossil is part of the Lower Cambrian and is no doubt the oldest
representative of modern crustaceans (
Link
).
Crayfish
were once thought to have originated about 140 million ears ago (compare
with the oldest known lobster at 167 Ma;
Link). Most researchers assumed that crayfish had descended from
lobsters around that time and gradually made their way inland. However,
Hasiotis has discovered 220-million-year-old specimens that are almost
identical to modem ones. He thinks crayfish may be as much as 300 million
years old. (
Link
).
There is also the problem of last appearance, which is becoming similar to the problem of first appearance. Mike Benton, Professor of Vertebrate Paleontology at the University of Bristol, noted an interesting phenomenon along these lines when studying the KT-boundary. During a program entitled, "The KT Boundary"(June 23, 2005), as part of the BBC radio series In Our Time, Benton commented:
"The fact is that many major groups of organisms did die out at that time, rather catastrophically, and I've observed that over the last 10 or so years as people have looked at the fossil records ever more closely, certain records, for example the ammonites, this major group of marine organisms, used to look rather gradualistic, meaning species disappeared step by step, one at a time for many metres below the boundary, but when people have gone back to the sections and collected more intensely, they're filling the gaps, so a lot of these records that used to look gradual stepwise dropping off, become more and more catastrophic with time and more and more species go up to the boundary, and that's true of the dinosaurs as well." (Link)
Consider also that
the generally orderly distribution of fossils in the fossil record is also
somewhat problematic for those who hold that the geologic column and fossil
record developed over many hundreds of millions of years. It seems like
the fossil record is simply too neatly sorted. Creatures suddenly appear
without any prior record and then suddenly disappear without any subsequent
record, only to reappear tens of millions of years later - alive and well (such
as the history of the Coelacanth
described in more detail below). Many layers also have only a limited
number of preserved creatures - far too limited an array to support a viable
ecosystem. For example, the fossils of some layers consist primarily of
large meat eating dinosaurs. Yet, there simply aren't near the number of
preserved creatures that could serve as prey for these very large carnivores.1
What on Earth did they live on for millions of years? How did they get
preserved in abundance in certain layers while other creatures did not? The same
thing is true of millions of fossils of very large plant eating dinosaurs in
places like the Morrison Formation. This
huge formation contains millions of preserved plant eating dinosaur bones but
hardly any plant fossils.34,35 Could the difference in
preservation between dinosaur bones and hard woody plant remains be that
significant? - especially when large seems of coal and other finely preserved
plant fossils are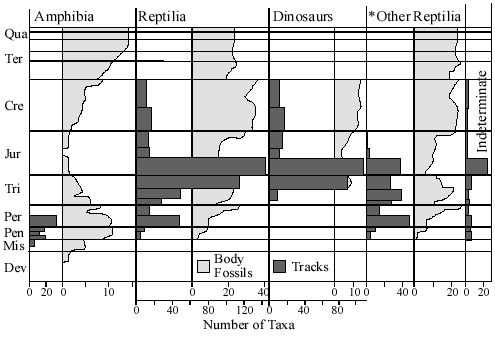 abundant elsewhere? And, what about pollen
order? Pollen appears fairly neatly sorted in the fossil record.
It seems, perhaps, that it is just too sorted.
abundant elsewhere? And, what about pollen
order? Pollen appears fairly neatly sorted in the fossil record.
It seems, perhaps, that it is just too sorted.
There are so many other features of the geologic column and fossil record that seem just as difficult, if not more so, for the notion that very long periods of time are represented. For example, it seems that many land animals, excluding birds and mammals, do not generally have their footprints located in the same layer in which their bodies are found, but in lower layers.56 Did the footprints evolve before they did? The footprints of dinosaurs, for example, are generally located in lower levels than the actual fossilized bones of the dinosaurs.1,56,82 Why would this be? What is there to explain this apparent sorting of body from footprint fossils? Leonard Brand and James Florence comment on this most interesting phenomenon:
If the geologic column represents sediments that have accumulated over many millions of years, and the fossils from each geologic period are the remains of animals living in successive time periods, it would be reasonable to expect that the stratigraphic patterns of footprint diversity should roughly parallel the patterns of equivalent body fossil diversity.56
Some have suggested various potential problems for this interpretation of Brand and Florence. However, these objections seem fairly well covered in the paper. I also discuss a few of these objections in detail in a Google Talk.Origins debate (Link).82
Some argue that such finely preserved trackways as are observed in
the fossil record, such as the horseshoe crab fossil and trackway pictured here,57
were probably the result of the creature being trapped in an "anoxic lagoon".
But how are such crisp footprints and such a well-preserved body going to
be preserved at the milky bottom of some anoxic lagoon?
Where are such conditions described preserving such fine fossils today?
Also, scavengers are still active even in highly anoxic conditions.
What made it possible for such delicate footprints and bodies of such
creatures to be preserved in such fine detail - avoiding all hints of
bioturbation?
Bioturbation is an extremely effective way of destroying layering in sedimentary rocks by mixing up the sediment and homogenizing it. It is easy to find modern-day examples of this. Hurricane Carla laid down a distinctive layer of sediment off the coast of central Texas in 1961. About twenty years later, geologists returned to find out what had happened to this layer. Most of the layer had been destroyed by living creatures burrowing into it and disturbing it; and where the layer could still be found it was almost unrecognizable.
In the light of such modern day findings, it is very difficult to imagine how such layering of sediment found throughout the geologic column and such crisp lines between these layers could have been kept in such pristine condition for not only tens or hundreds of years, but hundreds of thousands and even millions upon millions of years of time. It is even more difficult for me to imagine how such finely detailed fossils and trace fossils could be preserved.
Rather, it seems to me that the fossil presented here, with its trackway, was preserved by rather rapid and deep burial by sedimentary layering. The horseshoe crab survived the first periods of layering, making its trackway on a newly deposited surface, only to be overcome quite suddenly by a subsequent depositional events which were so rapid that the horseshoe crab, which was probably very good at digging, was trapped with such pressure that it could not escape. Its body and fine trackways were also preserved because of the sheer deepness of the burial - which prevented subsequent disturbance by bioturbation.
To suggest that such trackways, in particular, could be preserved in the bottom of some anoxic lagoon just doesn't make nearly as much sense to me. However, if someone can show me a real life example, I'd be more open to such potential explanations.
Also, what is usually overlooked is that
fact that there is no such animal as a "simple" animal. All living things
are extraordinarily complex. The "simplest" one celled organism, such as a
bacterium, is just a complicated as a single cell that exists in the body of a
human being - using just as many genetic functions at any one time. A rat
is also no less complex than a human as far as the information needed to build a
rat. In fact, out of 35,000 to 40,000 genes that the human genome
contains, the rat is only about 500 genes different.9 Certain
plant species also require a similar amount of functional genetic material
(i.e., genes). 
Consider also the fact that many mainstream scientists believe that almost every metazoan phylum with hard parts, and many that lack hard parts, made a first appearance in the Cambrian. The only modern phylum with an adequate fossil record to appear after the Cambrian was the phylum Bryozoa, which was not known before the early Ordovician until 2010 when Landing et. al. discovered a late Cambrian specimen in southern Mexico. 110
Of course, not all animal phyla have actually been found in the Cambrian layer since many have only soft body parts that are difficult to preserve or fossilize. Still, based on phylogenic analysis nearly all animal phyla that exist today are believed by most scientists to have arisen during or just before the Cambrian period over the course of a few tens of million years.
This rather sudden appearance, relatively speaking, of essentially all animal phyla during the Cambrian period is referred to as the 'Cambrian Explosion'. After this period essentially no new phyla are thought to have evolved over the course of hundreds of millions of years. Compared with the 35 or so animal phyla that still exist today, some people estimate that the Cambrian explosion may have generated as many as 100 different phyla. This prompted those such as Roger Lewin to ask, "Why, in subsequent periods of great evolutionary activity when countless species, genera, and families arose, have there been no new animal body plans produced, no new phyla?" 78 (Back to Top)
 As far as the fossilized bones of large animals, such as the
dinosaurs and large mammals, they are also generally oriented in the same
direction for any given layer, and this is true the world over.
Did these animals position themselves in the same direction as they died?
This does not really sound too likely. Even the legs and
tails of these animals are oriented in the same direction for a given
sedimentary layer. How does this happen? If you think
this is strange, consider also that huge masses of large bones are found matted
together in places like Bighorn Wyoming. Did these animals choose to die
in the same location and in the same general orientation? Some have argued that
the bones from Bighorn, Wyoming (pictured to the right) are not really oriented
since they obviously pictured as pointing in different directions. However,
water flow orients long thin objects in two ways - perpendicular and parallel
relative to the flow of the water. There are many places where
literally thousands of fossilized skeletons can be found all mixed up together
As far as the fossilized bones of large animals, such as the
dinosaurs and large mammals, they are also generally oriented in the same
direction for any given layer, and this is true the world over.
Did these animals position themselves in the same direction as they died?
This does not really sound too likely. Even the legs and
tails of these animals are oriented in the same direction for a given
sedimentary layer. How does this happen? If you think
this is strange, consider also that huge masses of large bones are found matted
together in places like Bighorn Wyoming. Did these animals choose to die
in the same location and in the same general orientation? Some have argued that
the bones from Bighorn, Wyoming (pictured to the right) are not really oriented
since they obviously pictured as pointing in different directions. However,
water flow orients long thin objects in two ways - perpendicular and parallel
relative to the flow of the water. There are many places where
literally thousands of fossilized skeletons can be found all mixed up together  in mass burial. In many of these places the bones
are severely damaged, fractured, and mangled - yet oriented.
It is all quite interesting as well as very difficult for today's popular
scientist to easily explain, if at all, without using a catastrophic flood
model.1
in mass burial. In many of these places the bones
are severely damaged, fractured, and mangled - yet oriented.
It is all quite interesting as well as very difficult for today's popular
scientist to easily explain, if at all, without using a catastrophic flood
model.1
Consider also the Jurassic Morrison Formation (famous for its dinosaur fossils). It covers over 1,000,000 square kilometers - being spread from Canada to Texas. It has been suggested that it was distributed by widespread flowing water. The fossils found within it, millions upon millions of them, are generally oriented with respect to flow - confirmed by GPS mapping (Arthur Chadwick). However, ancient channels of major rivers that would help distribute the sediments over such a wide area have not been found. Jack Horner noted the same thing, orientation with respect to flow, in the Montana deposits containing tens of thousands of dinosaurs.58 (Back to Top)
The Exquisite Preservation of Large Fossils
 The bodies of some very large fossils, such as dinosaurs or whales
are occasionally very well preserved. Of course, the geologic
layers that contain their fossilized remains are supposed to represent thousands
or even millions of years of Earth's history. However, the bodies of these
large creatures take up a fair percentage of the thickness of some of these
layers. This might just pose a bit of a problem for the
standard way of interpreting these fossils and the layers of sediment in which
they are found. Obviously, If
sedimentation slowly buried them over the course of very long periods of time,
their bodies would not have survived. For
fossilization to occur, burial or some other form of preservation must be fairly
rapid in order to protect the remains from significant scavenging and/or decay.
The bodies of some very large fossils, such as dinosaurs or whales
are occasionally very well preserved. Of course, the geologic
layers that contain their fossilized remains are supposed to represent thousands
or even millions of years of Earth's history. However, the bodies of these
large creatures take up a fair percentage of the thickness of some of these
layers. This might just pose a bit of a problem for the
standard way of interpreting these fossils and the layers of sediment in which
they are found. Obviously, If
sedimentation slowly buried them over the course of very long periods of time,
their bodies would not have survived. For
fossilization to occur, burial or some other form of preservation must be fairly
rapid in order to protect the remains from significant scavenging and/or decay.
With this in mind, it seems like the entire layer that such fossils are found in could not possibly have been formed over vast periods of time. Rather, it seems like such fossils speak of a relatively sudden burial event or events, or some sort of rapid process of fossilization without burial - sometimes on massively catastrophic proportions covering hundreds of thousands or even millions of square kilometers. The very high degree of preservation of fine details of some of these fossils give evidence of a rather sudden burial or preservation process, and not of a natural death with slow burial or preservation giving time for decay. Many fossils show evidence of surprise or being startled, such as elevated dorsal fin spines or tightly close clams, or a brief struggle before death - like they were suddenly buried or rapidly preserved by other processes (like sudden crystallization) extremely quickly. Some fossils are found with food still in the mouth of the victim (in mid chew). Others have been found suddenly frozen in the process of giving birth and others have been found with fine soft tissue detail down to the cellular level (still turgid gills of fish preserved in the Santana Formation - indicating complete fossilization in less than 1 hour).
It is my personal opinion that such fossil evidence favors a catastrophic interpretation for much of the geologic column. Interestingly, this notion isn't as "fringe" as it used to be just a few decades ago. Mainstream scientists are starting to lean more and more toward catastrophism. This is quite interesting because, for a very long time, mainstream scientists where "uniformitarian" in their thinking.
"From around 1850 to 1980, most geologists endorsed uniformitarianism ("The present is the key to the past") and gradualism (geologic change occurs slowly over long periods of time) and rejected the idea that cataclysmic events such as earthquakes and volcanic eruptions played any significant role in the formation of the Earth's surface. In part, the geologists' rejection was fostered by their impression that the catastrophists of the nineteenth century believed that God was directly involved in determining the history of Earth. Catastrophism of the nineteenth and early twentieth centuries was closely tied to religion and catastrophic origins were considered miraculous rather than natural events."79
The uniformitarian notion of slow deposition over millions of years of time is no longer accepted by scientists. It is now believed that various catastrophes, large and small and sometimes worldwide, played a key role in the Earth's history and are recorded in the geologic column. In this regard, consider the following comments from David, M. Raup of the Chicago Field Museum of Natural History:
"A great deal has changed, however, and contemporary geologists and paleontologists now generally accept catastrophe as a 'way of life' although they may avoid the word catastrophe... The periods of relative quiet contribute only a small part of the record. The days are almost gone when a geologist looks at such a sequence, measures its thickness, estimates the total amount of elapsed time, and then divides one by the other to compute the rate of deposition in centimeters per thousand years. The nineteenth century idea of uniformitarianism and gradualism still exist in popular treatments of geology, in some museum exhibits, and in lower level textbooks....one can hardly blame the creationists for having the idea that the conventional wisdom in geology is still a noncatastrophic one."80
Also, consider the following comments by Robert H. Dott
given during a Presidential Address To Society of Economic
Paleontologists & Mineralogists:
"I hope I have convinced you that the sedimentary record is largely a record of episodic events rather than being uniformly continuous. My message is that episodicity is the rule, not the exception. . . We need to shed those lingering subconscious constraints of old uniformitarian thinking."81
It seems then that catastrophism is only recently being accepted by more mainstream scientists. It seems also that significant portions of many different sedimentary layers within the geologic column are now thought, even by popular science, to have been the result of rather sudden catastrophic deposition - with the bulk of time passing in between these episodes of catastrophe. Although the notion that the geologic column might not actually represent millions of years of time is far from mainstream, mainstream thinking is actually drifting back toward a position that can actually begin to consider that catastrophes and episodicity are "the rule, not the exception." This notion is something fairly new in mainstream thinking. Even today there is strong resistance of any notion that comes to close to suggesting catastrophes of "Biblical proportions" - perhaps due to the engrained bias against literal Biblical interpretations that suggest such things as a worldwide flood. Consider the story of J Harlen Bretz as a fascinating example of this sort of resistance.
In this line, the fairly recent discoveries of fossil whales (Miocene/Pliocene) in western Peru are quite interesting. Leonard Brand (Ph.D. in Paleobiology from Cornell) comments, "In our survey of the area, we found the fossil remains of more than 100 whales in an area of less than two square kilometers. What was even more exciting was the well-preserved nature of the fossil remains. . . Typically, when a whale dies at sea, the carcass falls to the bottom and becomes the source of a rich ecosystem. Many species of sea life benefit from the decaying remains at each stage of the process. Within four to six months, the whale carcass has been mostly stripped down to the bones. At that point, other species of organisms burrow both into the bones and the surrounding sediment. Within a year or two, the whale bones show much evidence of these burrowing animals."52
So, how did the whales in western Peru meet their end? "These whales were incredibly well-preserved," Brand observes, "suggesting that they were covered quickly." Brand found that the whale remains were blanketed by a thick layer of diatomite (silica remains of diatoms). These tiny creatures, known collectively as plankton together with dinoflagellates, are part of the food source for whales. In modern times, diatomite normally accumulates on the sea bottom at a rate of a few centimeters per thousand years. "We also found beautifully preserved baleen," he adds. Baleen refers to the filtering feather-like structures in the whale's mouth that are used to strain out food (plankton) from the water. "Whales feed by gulping in water and forcing it out through the baleen, trapping the tiny plankton." Baleen is actually more akin to the human fingernail or toenail in its structure. "The well-preserved baleen supports the theory of a quick burial to an even greater extent".52
But why did the whales die in the first place? "There is more and more evidence that red tides--blooms of diatoms and dinoflagellates--produce toxins which can kill large animals and fish," he says.52 These massive blooms were so large that they not only killed the whales, but buried them in thick layers before any significant decay could set in.
 Another very startling finding that demonstrates the
sudden/catastrophic burial of very large creatures is a 1971 finding in Southern
Mongolia of a perfectly articulated Protoceratops and a Velociraptor frozen in a
life and death struggle with each other. Obviously
these two creatures were buried suddenly by a huge catastrophe of magnificent
proportions. The dinosaurs didn't even have time to
fall over. 51
Another very startling finding that demonstrates the
sudden/catastrophic burial of very large creatures is a 1971 finding in Southern
Mongolia of a perfectly articulated Protoceratops and a Velociraptor frozen in a
life and death struggle with each other. Obviously
these two creatures were buried suddenly by a huge catastrophe of magnificent
proportions. The dinosaurs didn't even have time to
fall over. 51
Many dolphin-like ichthyosaur fossils also show evidence of rapid
burial - such as those found clustered together at places like the
Berlin-Ichthyosaur State Park in Nevada. Some
ichthyosaurs that show evidence of rapid burial are buried with certain other
ichthyosaurs that show evidence of rather brief exposure and/or scavenging in
the same region.
Isn't it strange that creatures showing evidence of some exposure are buried
closely with others that show
 evidence of very rapid burial?
According to scientists studying these fossils, "It is not yet certain how these
large creatures died and were buried together in such a small area."
Also, at least in the Nevada location, "the skeletons are generally
oriented along a north-south axis, suggesting that currents or tides played some
part in deciding their final resting place."
evidence of very rapid burial?
According to scientists studying these fossils, "It is not yet certain how these
large creatures died and were buried together in such a small area."
Also, at least in the Nevada location, "the skeletons are generally
oriented along a north-south axis, suggesting that currents or tides played some
part in deciding their final resting place."
Other evidences of rapid ichthyosaur burial include some specimens
that were suddenly buried in the middle of giving birth!
Consider also the large size of adult ichthyosaurs.
In order to preserve such large specimens, in such well-preserved
condition, fairly rapid burial is  required.
These are all indications of some sort of sudden event that resulted in
the relatively simultaneous deaths of many ichthyosaurs as well as their
relatively rapid burial.
required.
These are all indications of some sort of sudden event that resulted in
the relatively simultaneous deaths of many ichthyosaurs as well as their
relatively rapid burial.
Some prolonged exposure of such large creatures as ichthyosaurs is only to be expected, but the fact that their fossil remains were preserved in an articulated fashion, often with evidence of soft tissue detail, speaks in favor of a fairly rapid burial process with little chance for significant decay and/or scavenging. Also, many creatures may die at the same time and then be buried rapidly at slightly different times by repeated waves of sediment deposition.53 (Back to Top)
Peru's Fossil Whales Challenge Radiometric Dating Assumptions
In 1999 Dr. Raul Esperante teamed up with Dr. Leonard Brand and others to investigate fossil whales within the Pisco Formation of Peru's Atacama Desert. This formation is approximately 600 meters thick and consists of many layers of sedimentary rock. It is bounded by two layers of volcanic ash with the lower ash layer dating 12 million years older than the upper ash layer (dated by potassium-argon; K/Ar). This means that, in standard geological thinking, the 600 meters of sedimentary rock between the ash layers must have been deposited over the course of some 12 million years of time (~20,000 years per meter). Yet, within essentially all of these layers are hundreds of very well preserved fossil whales. In fact, many of them are so well preserved that their baleen is still intact and attached in the usual position that baleen is attached in living whales. Usually baleen detaches within a few days (or even hours) after death. Some of the fossilized whales and dolphins also have preserved remains of skin outlines around the fossilized bones. The skeletons themselves are generally well articulated and show no evidence of scavenging or significant decay.
There are several problems that these fossil whales pose for mainstream assumptions regarding radiometric dating since these features are more consistent with a catastrophic/rapid formation of all of the fossil-bearing layers within a much much shorter period of time than radiometric dating suggests:
The fossil whales must have died and been completely buried by diatomaceous sediment within a very short time of death (no scavenging, decay, significant disarticulation, or loss of baleen).
The layers are very smooth without significant erosion or unevenness to suggest the passage of time between layers.
There is no significant bioturbation (very few tunnels or evidence of trace fossils or digging within the sedimentary layers) that would be expected given long periods of time between the formation of subsequent layers.
There are finely preserved shards of volcanic glass within all of the layers that have very sharp edges without the usual rounding that would be expected (due to the relatively rapid ability of water to dissolve silica) if long periods of time took place during the build up of these sedimentary layers.
These layers were deposited in shallow seas with evidence of flowing currents, which works against the potential counter-hypothesis that these layers were formed under anoxic conditions.
The Living Fossils

The coelacanths are fish having a flexible oil-filled type of back bone that gives them their name. In place of the bony vertebral column of most adult fishes, coelacanths have a large, thick tube of cartilage, called the 'notochord'. In the early development of most fishes, the notochord of the embryo or larva is gradually replaced by the bony (or calcificed) center of the vertebral column. But in coelacanths, lungfishes and some types of sharks, the transformation of notochord into a segmented bony (or calcified) vertebral column does not take place. In coelacanths, the hollow notochord is filled with oil and provides a strong, yet flexible support for the spinal cord. 41
 Coelacanths are thought to be very ancient fish who lived
before, during, and after the time of the dinosaurs. However, like the
dinosaurs, the coelacanths vanished from the fossil record (some 80 million
years ago). However, before they seemingly went extinct they lived
prosperously for nearly 400 million years. They are in fact thought to be
the ancestors to the first vertebrates to step out of the seas and walk on dry
land some 350 million years ago. Because of this belief that coelacanths
are an important evolutionary link between sea and land dwelling vertebrates,
special significance has been given to their place in the fossil record.
In fact, it was long thought that the fossil record was the only place where
coelacanths would ever be found. This seemed to be true until 1938 when
living coelacanths were found alive and well off the coast of South Africa
living at a depth of around 120 to 250 meters. 41
Since then, other colonies of coelacanths have been found in various areas such
the coast of Madagascar, Sulawesi (Indonesian Archipelago), Mozambique, and
Comoran.
Coelacanths are thought to be very ancient fish who lived
before, during, and after the time of the dinosaurs. However, like the
dinosaurs, the coelacanths vanished from the fossil record (some 80 million
years ago). However, before they seemingly went extinct they lived
prosperously for nearly 400 million years. They are in fact thought to be
the ancestors to the first vertebrates to step out of the seas and walk on dry
land some 350 million years ago. Because of this belief that coelacanths
are an important evolutionary link between sea and land dwelling vertebrates,
special significance has been given to their place in the fossil record.
In fact, it was long thought that the fossil record was the only place where
coelacanths would ever be found. This seemed to be true until 1938 when
living coelacanths were found alive and well off the coast of South Africa
living at a depth of around 120 to 250 meters. 41
Since then, other colonies of coelacanths have been found in various areas such
the coast of Madagascar, Sulawesi (Indonesian Archipelago), Mozambique, and
Comoran.
The initial discovery of living coelacanths came as quite a shock to the scientific community. To find a "living fossil" after it had vanished from the fossil record for some 80 million years was quite a stunning revelation. How could this be explained? How could a creature be preserved/fossilized over and over again during the course of 300+ million years and then vanish for some 80 million years only to reappear, alive and well, in modern oceans?
Well, there are several explanations commonly given to explain such a phenomenon. Perhaps one of the more popular explanations is that the modern coelacanth is not the same creature as those coelacanths found in the fossil record. In fact, the differences are thought to be so significant that modern coelacanths have not only been classed in a different species category, but in a different genus group as well. The classification of the modern coelacanth Latimeria chalumnae verses the very similar fossil coelacanth Macropoma lewesiensis, goes as follows:
|
|
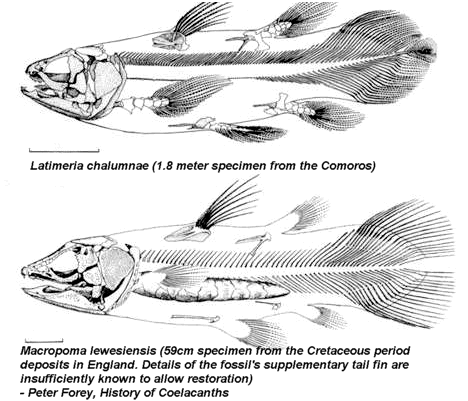 Many "emphasize" the point that, "The living coelacanth is not a living fossil
in the very strict sense that members of the species L. chaumnae itself
have never been found as a fossil. In fact, no other species assignable to
the Genus Latimeria has been found as a fossil either. Latimeria
and the Cretaceous fossil Genus Macropoma
are quite closely related, and we could possibly include them in the same
family. Beyond that, all fossil coelacanths belong to the order
Coelacanthini."
38,40
Many "emphasize" the point that, "The living coelacanth is not a living fossil
in the very strict sense that members of the species L. chaumnae itself
have never been found as a fossil. In fact, no other species assignable to
the Genus Latimeria has been found as a fossil either. Latimeria
and the Cretaceous fossil Genus Macropoma
are quite closely related, and we could possibly include them in the same
family. Beyond that, all fossil coelacanths belong to the order
Coelacanthini."
38,40
The problem with this argument, as discussed in some detail above, is that phylogenic classification schemes are quite subjective. A comparison of the modern coelacanth with those found in the fossil record really does not necessitate their being any more different from various breeds of dogs who are all members of not only the same genus and family groups, but the same species group as well. Even so, some do argue that, "A person can see substantial differences in the fins, tail, lobe fin, and drastic changes in the structure of the head." 40 However, it seems to me that this is an over emphasis when such differences are no more drastic than the differences that are seen within species groups (ie: bulldog vs. German Shepherd). It is true that the picture comparisons do show some obvious differences, but these do not seem to be as striking as many seem to suggest. The lobe fins themselves are almost identical. The tails are also practically identical excepting for comparisons of the "supplementary tail fin." The problem here is that the supplementary tail fin was left out of the drawing of the fossil coelacanth, Macropoma lewesiensis (as were the fin bones). This is because, "Details of the fossil's supplementary tail fin are insufficiently known to allow restoration" (The same can be said for the fin bones). This leaves, basically, the "drastic changes in the structure of the head" as the only real possible difference between the two coelacanths.
It appears that the two heads are in fact very similar as far as the number, relative position, and general shape of the bones in the two creatures. It seems like the main differences can be found in the relative size of the various bones or bone proportions. These differences give Macropoma a longer and more pointed snout than is found in Latimeria as well as a bit of an overbite.
Based on these rather minor differences these two creatures are classed, not just as different species, but as members of different genus groups. Are these differences really greater than the range of comparable differences found within the range of other species, such as domestic dogs or even modern humans? Consider, for example, the differences between a German Shepherd and a bulldog. The German Shepherd has a long, narrow snout with an overbite, while a bulldog has a short, flat snout with an underbite. These differences might even be called "drastic." If the bulldog phenotype where found only in the fossil record and the German Shepherd were living today, would they be classed in the same species or even genus groups when compared side-by-side? I doubt it.
In this light, the statement that "No other species assignable to the Genus Latimeria has been found as a fossil", seems to me to be a bit misleading. In fact, Peter Forey, in his book, "History of the Coelacanth Fishes", comments that the skeleton of Macropoma lewesiensis is "virtually identical to that of the coelacanths caught off the Sodwana Bay, Latimeria chalumnae, and differs little from the skeleton of most Devonian coelacanths." 39,40 The problem is that there does in fact seem to be a tendency to place very similar fossils in different categories based primarily on the fact that they obviously lived so far apart in time that they can not possibly be members of the same species. This practice seems to be the rule rather than the exception. So, to say that the L. chaumnae species has never been found in the fossil record seems to me to be stretching it a bit since this species classification is based on very minor morphologic differences that are clearly within the range of intra-species variation.
So why then, did the coelacanth disappear from the fossil record for 80 million years? Other arguments are that only those coelacanths that lived in more shallow water environments died out while the deep water variety managed to survive. The deep water environment occupied by many modern coelacanths is thought to be very poorly conducive to both fossilization and discovery. Other coelacanths, such as those colonies that have been found more recently living off the coast of the Comoro Islands, live in shallower waters but over steep slopes of these volcanic islands. Some scientists, such as Hans Fricke (a coelacanth specialist) suggest that, "these slopes are too steep to hold sediments that could rapidly cover a dead fish and thus begin the fossilization process." The steep slopes and underwater caves that form the habitat for the modern coelacanth simply do not allow for the fairly rapid sedimentation and burial needed for fossilization to occur. Also, deep water prevents scientists from exploration and discovery even if such fossilization did occur. Additionally it is thought that continental subduction might have destroyed such deep sea fossils as well.40
These arguments are all well and good except for one thing. If coelacanths were so successful for hundreds of millions of years in areas quite conducive to successful fossilization, then why, when they died out of such areas, did they not evolve back into these niches over the course of time? Why did these niches that supported coelacanths so successfully over the course of hundreds of millions of years never become repopulated over the course of 80 million years by populations of coelacanths living elsewhere? Really, 80 million years is a fair amount of time. One might think that surviving populations of deep-sea coelacanths might fluctuate in size. They might even "evolve" the ability to populate rivers and lakes again. Is it not strange that in 80 million years, the surviving populations of coelacanths never found opportunity to evolve back the ability to live in their former habitats where they were previously so successful? After all, it seems like a much shorter step to go from a deep sea environment to a shallow water environment than to go from swimming to walking around on land. If the coelacanths gave rise to land dwelling creatures in a few million years, then why was it so difficult for them to win back their former water-based environments? (Back to Top)
Dinosaur Eggs
Added September, 2011

 There
are many who argue that dinosaur eggs clearly falsify the
concept of a recent global Flood of Noachian proportions.
However, as far as I've been able to tell, it seems like
dinosaur eggs actually lend greater support to the worldwide
Flood model. I certainly don't see how dinosaur eggs
definitively undermine this model as many suggest.
There
are many who argue that dinosaur eggs clearly falsify the
concept of a recent global Flood of Noachian proportions.
However, as far as I've been able to tell, it seems like
dinosaur eggs actually lend greater support to the worldwide
Flood model. I certainly don't see how dinosaur eggs
definitively undermine this model as many suggest.
Consider, for example, certain general features of dinosaur
eggs:
Of the hundreds of thousands of eggs that have been preserved in
the fossil record it seems likely that over 99% of them contain
no embryo.
Essentially all of the eggs that have been
found were buried by water-born sediments around the world.
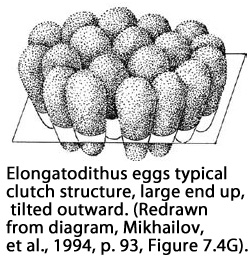
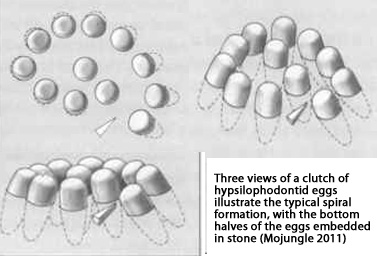
Many examples of egg beds were laid as
sediments were being actively deposited - to include
striking examples of eggs within the same "nest" being deposited
on multiple levels of sediment (see picture above).
 Most
asymmetrical eggs (eggs with a pointy end) were unexpectedly
preserved with the pointy end pointed downward and the larger
end pointed upward with a symmetrical inward or outward leaning
orientation - consistent with being laid in semi-liquid sediment
(like very watery mud).
Most
asymmetrical eggs (eggs with a pointy end) were unexpectedly
preserved with the pointy end pointed downward and the larger
end pointed upward with a symmetrical inward or outward leaning
orientation - consistent with being laid in semi-liquid sediment
(like very watery mud).
Those eggs that are found with "hatch windows" often contain the
shell fragments from the window within the egg itself - a
feature not expected from hatched eggs where the shell fragments
should be on the outside of the egg following hatching.
The overall arrangement of eggs in a nest with
"hatch windows" is not disturbed as would be expected from the
hatchlings moving the light eggshells around after hatching from
their eggs.
 Trackways
of young or baby dinosaurs are extremely rare relative to adult
trackways and the trackways that are found (of the adults) are
generally found in lower sedimentary layers compared to the body
fossils (Leonard Brand).
Trackways
of young or baby dinosaurs are extremely rare relative to adult
trackways and the trackways that are found (of the adults) are
generally found in lower sedimentary layers compared to the body
fossils (Leonard Brand).
Extremely well preserved embryos from Auca
Mahuevo (Argentina), to include the preservation of very
delicate embryonic bones and skin, suggests very rapid burial in
a supersaturated watery environment.
Commonly identified double layered egg shells suggesting the existence of a stressful environment worldwide (see discussion below).
Now, I do recognize that embryos, though very rare, are sometimes identified
(both within and outside of their eggs). I'm also aware that they show
fairly advanced development, to include fully formed skeletons and occasional
teeth. However, this is not entirely unexpected given the Biblical
Flood model (on a worldwide scale). As originally proposed by Leonard
Brand (Link),
cases are known were reptiles, like the Komodo Dragon for example, will withhold
the laying of fertilized eggs until a more favorable opportunity arises or until
they are put into very stressful conditions of "fight or flight". Of
course, if eggs are held for too long before being laid, they will develop a
"second shell" which suffocates the embryo. Dinosaur eggs have often been
found with such a double shell, suggesting that they were able to avoid laying
their eggs for some time in the hopes of more favorable conditions.
 Healthy,
well-adjusted chickens and marine turtles have only one shell round their eggs.
Give chickens a hard time, says Sally Solomon of Glasgow university, and they
react in a tell-tale way.
Healthy,
well-adjusted chickens and marine turtles have only one shell round their eggs.
Give chickens a hard time, says Sally Solomon of Glasgow university, and they
react in a tell-tale way.
"They retain the eggs in the reproductive tract and, in retaining it, it either gets an extra coating of calcium or sometimes it actually shoots back up the reproductive tract and it gets an extra layer of shell," she says. "If you take a busload of tourists on to a beach when turtles are trying to come ashore to lay their clutches of eggs in the sand, they will abandon the process and move back into the sea. When they are in the sea, the eggs are held and an extra layer of calcium is laid down. So you end up with a very thick shell."
What was true for birds and reptiles today must have been true for the ancestors of both birds and reptiles. When Frankie Johnson, a paleontologist working in Montana, sent her samples of eggs from fossil nests of Troodon, Professor Solomon recognized the symptoms immediately. The shells were double.
 "It's
a huge step to say it's stress in dinosaurs. But they must have had it, what
with their world collapsing about them. Here we have a phenomenon common to
dinosaurs, extant reptiles and birds. And we know for a fact that stress is
instrumental in causing double shelled eggs in turtles, poultry and many wild
birds. Is it too big a step to suggest that dinosaurs, despite their size,
also experienced stress? Those shells are abnormal: they were retained in the
oviduct for longer than normal. Why? What was there in that environment
which was inclement? These are questions we are looking at." 113
"It's
a huge step to say it's stress in dinosaurs. But they must have had it, what
with their world collapsing about them. Here we have a phenomenon common to
dinosaurs, extant reptiles and birds. And we know for a fact that stress is
instrumental in causing double shelled eggs in turtles, poultry and many wild
birds. Is it too big a step to suggest that dinosaurs, despite their size,
also experienced stress? Those shells are abnormal: they were retained in the
oviduct for longer than normal. Why? What was there in that environment
which was inclement? These are questions we are looking at." 113
I've read counter arguments suggesting that dinosaurs were warm blooded and
therefore could not retain their eggs for very long, but this is debatable and
is essentially falsified by the fairly common finding of thickened or doubled
dinosaur egg shells within the fossil record.
I've also read arguments where the suggestion is made that dinosaurs likely laid
only one or two eggs per day. Yet, this is clearly mistaken in many cases
where there is evidence of many eggs being laid in a very symmetrical pattern
within the same day. Also, I don't agree with the concept that the Flood
would have had to kill off all the dinosaurs by the 40th day. I think that
the Flood was complex, not a uniform increase of water over the globe. It
seems to me at least possible that animals could have survived the initial
months of the Flood - perhaps close to 150 days from the beginning of the Flood.
Noah was on the Ark, after all, for over a year.
(Back to Top)
Feathered Dinosaurs
Added April 10, 2012

Modern scientists have long believed and taught that modern featured birds originally evolved from theropod dinosaurs. After all, birds and theropod dinosaurs, or reptiles in general, share many of the same or similar features. For example, like all other reptiles, birds have scales. Also, birds lay eggs as do reptiles. Certain anatomical features, such as the overall musculatures, brain, heart, and other organs, as well as the skeletal system, are fairly similar.
In short, ancient birds shared the following major skeletal characteristics with many certain dinosaurs (especially the Maniraptora, which includes Velociraptors):
Pubis (one of the three bones making up the vertebrate pelvis) shifted from an anterior to a more posterior orientation, and bearing a small distal "boot".
Elongated arms and forelimbs and clawed manus (hands).
Large orbits (eye openings in the skull).
Flexible wrist with a semi-lunate carpal (wrist bone).
Hollow, thin-walled bones.
3-fingered opposable grasping manus (hand), 4-toed pes (foot); but supported by 3 main toes.
Reduced, posteriorly stiffened tail.
Elongated metatarsals (bones of the feet between the ankle and toes).
S-shaped curved neck.
Erect, digitgrade (ankle held well off the ground) stance with feet postitioned directly below the body.
Similar eggshell microstructure.
Teeth with a constriction between the root and the crown.
Functional basis for wing power stroke present in arms and pectoral girdle (during motion, the arms were swung down and forward, then up and backwards, describing a "figure-eight" when viewed laterally).
Expanded pneumatic sinuses in the skull.
Five or more vertebrae incorporated into the sacrum (hip).
Straplike scapula (shoulder blade).
Clavicles (collarbone) fused to form a furcula (wishbone).
Hingelike ankle joint, with movement mostly restricted to the fore-aft plane.
Secondary bony palate (nostrils open posteriorly in throat).
Possibly feathers... this awaits more study. Small, possibly feathered dinosaurs were recently found in China. It appears that many coelurosaurs were cloaked in an external fibrous covering that could be called "protofeathers."
Such similarities have long been recognized. Many anatomists between the 1500s to 1800s noticed that birds shared various similarities to reptiles. Then, in 1860, the first specimen of Archaeopteryx lithographica (pictured above) was discovered by a quarry worker in Germany. For many, it was a beautiful example of a "transitional form" between reptiles and birds - and seemed to confirm Darwin's recently published expectations. J. H. Ostrom's 1969 description of Deinonychus antirrhopus and its similarities to Archaeopteryx then provided the groundwork for the modern view of birds evolving the ability to fly with the use of feathers. Then, Dr. Gautheir's cladistic work in the mid-1980's provided analytical systematic support for the dinosaur-bird evolution theory.
Of course, there has been some disagreement among modern scientists as to the
true evolutionary origin of birds. Dr. Alan Feduccia, for example, has
been an outspoken opponent of the dinosaur-bird evolution hypothesis.
During his 2004 talk at the San Diego History Museum on the origin of birds, he
said, "Paleontologists
have tried to turn
Archaeopteryx
into an earth-bound, feathered dinosaur. But it's not. It is a bird, a perching
bird. And no amount of 'paleobabble' is going to change that." (Link)
During this talk he especially emphasized the following points:
Archaeopteryx is a true bird.
"Dinofuzz" is nothing more than collagenic fibers found on many
other fossils.
Today's highly touted "Feathered Dinosaurs" are a myth: some
fossils (i.e. Caudipteryx) have flight-feathers but they aren't
really dinos--they are secondarily flightless birds
Birds have digits 2-3-4, and theropods have digits 1-2-3. This
is powerful evidence that birds couldn't have evolved from
theropod dinos.
Also, the theropod --> bird hypothesis requires that birds
evolved flight from the ground-up. If
Caudipteryx
has feathers but not for flight, Feduccia finds this explanation
quite tenuous. Put simply, ground-up proponents say feathers
were pre-adapted for flight but evolved originally for
insulation. This is silly because feathers are perfectly suited
for flight, and very energetically costly to produce. If
insulation was all that was needed, hair would have done the job
just fine and would not have been nearly so costly. It strains
credibility to say feathers evolved for insulation.
Feduccia prefers Microraptor as an ancestor of birds because he
likes the trees-down hypothesis, not the ground-up hypothesis.
If birds didn't come from theropods, this does leave a rather
large time-gap where there is essentially no fossil
documentation of exactly what sort of dinos or other reptiles
from which birds would have evolved.
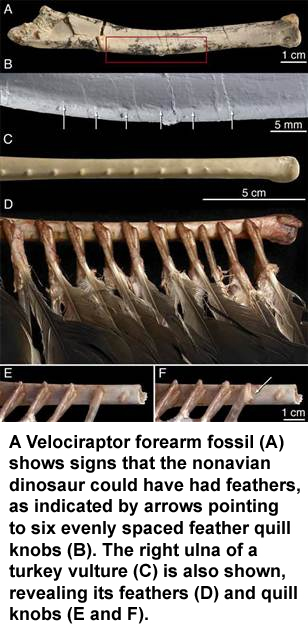
Of course, the fact that "quill knobs" have been shown on the arms of real dinosaurs like Velociraptors (pictured above) is quite difficult for many scientists to ignore. How can the pro and con arguments for key similarities and differences be resolved?
Well, some scientists are now arguing that dinosaurs evolved from birds, not the other way around. That's right, birds came first with their complex feathers and flight adaptations and then theropod dinosaurs lost these specialized features when they took to the ground.
“Raptors look quite a bit like dinosaurs but they have much more in common with birds than they do with other theropod dinosaurs such as Tyrannosaurus,” Ruben said. “We think the evidence is finally showing that these animals which are usually considered dinosaurs were actually descended from birds, not the other way around.”ScienceDaily (Feb. 9, 2010)
 So,
we have yet another example of devolution in action – the same mechanism that
produces flightless birds on windy islands or cavefish without eyes. This form
of change over time is very easy to explain since it is far easier to break
something via mindless mechanisms than it is to create a working complex system
to begin with via any known mindless mechanism.
So,
we have yet another example of devolution in action – the same mechanism that
produces flightless birds on windy islands or cavefish without eyes. This form
of change over time is very easy to explain since it is far easier to break
something via mindless mechanisms than it is to create a working complex system
to begin with via any known mindless mechanism.
Again, look as you might, you will not find an example in literature of evolution in action beyond very very low levels of functional complexity (i.e., beyond the level of 1000 specifically arranged amino acid residues), nor will you find a mathematical model that makes any useful predictions as to the success of the mechanism of RM/NS at various levels of functional complexity over a given period of time.
In short, faith in the RM/NS mechanism as the primary source of creativity in evolutionary biology for the production of higher level systems of function within gene pools is nothing but fairytale wishful thinking – not science. Functionally complex differences between different groups of animals, such as the complexity of true bird feathers, can only be explained by deliberate intelligent design.
 Shale beds, such as the Yesnaby Sea Stacks and the Dougherty Gap
Outcrop pictured here, are formations that are often composed of alternating
layers of shale and sandstone. The various layers range from one or two
millimeters to
Shale beds, such as the Yesnaby Sea Stacks and the Dougherty Gap
Outcrop pictured here, are formations that are often composed of alternating
layers of shale and sandstone. The various layers range from one or two
millimeters to  several meters in thickness. The layers of shale where
once layers of clay that have become compressed and hardened into shale.
It is generally thought that such layers were formed over several million years
by the repetitive deposits of shallow lakes, swamps, and rivers. With the
changing sea-levels due to glacial activity, the resulting cyclical drowning of
these areas is thought to have resulted in the cyclical deposition of clays,
silts and sands over fairly significant spans of time. Some of these beds
are in fact quite thick. For example, the Haymond Beds average around
1,300 meters in thickness and contain thousands of layers of shale and
sandstone. However, what is especially interesting about many of these
layered beds is that they contain "trace fossils".
59
several meters in thickness. The layers of shale where
once layers of clay that have become compressed and hardened into shale.
It is generally thought that such layers were formed over several million years
by the repetitive deposits of shallow lakes, swamps, and rivers. With the
changing sea-levels due to glacial activity, the resulting cyclical drowning of
these areas is thought to have resulted in the cyclical deposition of clays,
silts and sands over fairly significant spans of time. Some of these beds
are in fact quite thick. For example, the Haymond Beds average around
1,300 meters in thickness and contain thousands of layers of shale and
sandstone. However, what is especially interesting about many of these
layered beds is that they contain "trace fossils".
59
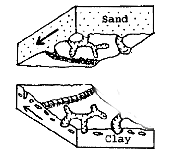
 Trace fossils are the evident remains of tracks or imprints that
some creature left behind even though the actual body is not there. For
example, when the shale was first formed as an organically rich clay, many
burrowing creatures lived in it, filtering it for nutrients. As they moved
through it, they left trails behind. When this clay was buried by
turbiditic sand flows, the sand filled in these tunnels, trails and other
impressions. As the sand solidified, the casts of these tunnels and other
markings were preserved on the underside of the sandy layer. Since this
underside of a layer is called the "sole", the preserved impressions in the clay
are called "sole casts."
Trace fossils are the evident remains of tracks or imprints that
some creature left behind even though the actual body is not there. For
example, when the shale was first formed as an organically rich clay, many
burrowing creatures lived in it, filtering it for nutrients. As they moved
through it, they left trails behind. When this clay was buried by
turbiditic sand flows, the sand filled in these tunnels, trails and other
impressions. As the sand solidified, the casts of these tunnels and other
markings were preserved on the underside of the sandy layer. Since this
underside of a layer is called the "sole", the preserved impressions in the clay
are called "sole casts."
 Many argue that these layers must have been formed over long
periods of time because colonies of such burrowing creatures take time to
colonize each layer of clay as it forms. The burrows themselves take a
fair amount of time to create. Glenn Morton, a vocal geologist, comments
that, "These burrows are horizontal and the animals don't seem to be digging
out. They are digging through the sediment. And there are thousands
of layers of sediment with the burrows on them." 61 Morton
actually suggests that when each sandy turbidite covered a layer of clay that
the burrowing creatures didn't burrow out, but died when the sandy layer covered
the layer of clay. He says, "We know that the burrowers who were buried
did not survive. If they had, they would have had to dig up through the
sand to escape their entombment. There are no burrowers going up through
the sand. And, if there had been these burrows, there should be little
circular piles of sand with a central crater pocking the entire upper surface of
the sand. We don't see these." 59
Many argue that these layers must have been formed over long
periods of time because colonies of such burrowing creatures take time to
colonize each layer of clay as it forms. The burrows themselves take a
fair amount of time to create. Glenn Morton, a vocal geologist, comments
that, "These burrows are horizontal and the animals don't seem to be digging
out. They are digging through the sediment. And there are thousands
of layers of sediment with the burrows on them." 61 Morton
actually suggests that when each sandy turbidite covered a layer of clay that
the burrowing creatures didn't burrow out, but died when the sandy layer covered
the layer of clay. He says, "We know that the burrowers who were buried
did not survive. If they had, they would have had to dig up through the
sand to escape their entombment. There are no burrowers going up through
the sand. And, if there had been these burrows, there should be little
circular piles of sand with a central crater pocking the entire upper surface of
the sand. We don't see these." 59
Glenn Morton is not the only one who thinks this way. This is in fact the prevailing paradigm about how these layers must have formed. However, there may be an even more reasonable explanation. If these layers were in fact formed over long periods of time where each individual layer took at least a few years to form, it seems like tunneling organisms would mess up the layers. Look at the pictures presented here. Most of these layers are very thin, averaging only a few centimeters in thickness. And yet, they are extremely crisp and distinct from the layers above and below. Burrowers living in lake or ocean bottoms or swampy areas, burrow all around and cause mixing of the sediments. Such mixing is known as "bioturbation." However, even the thinnest sandstone units fail to show any obvious signs of bioturbation, blurring of bedding contacts, or internal bedding features. Rather they appear as homogenized small-grained sandstones clearly demarcated from the overlying and underlying layers of shale. Further evidence suggesting a more rapid formation of the layers comes from work done by Kuenen in 1967. Kuenen documented the differences in sand textures between interdistributary bay deposits and turbidite deposits. Using his work as a reference the sandstone units found at Dougherty Gap best correlate to turbidite emplacement based on both lithology and bioturbation. Also, the work of Coleman and Prior gives even more support for this idea. In 1980 they presented photographs of cores taken from a modern interdistributary bay which in no way resemble the stratigraphy or sedimentation found exposed at the Dougherty Gap site. 60
Another interesting finding is that these layers get thicker as one move up the various outcrops. This finding is a common characteristic of rapid turbidite deposition and is "believed to reflect the progradation of submarine fan lobes." 60 In any case, this finding is not consistent with a slow cyclic deposition over vast spans of time.
The sand in the sand layers is also, "well sorted" meaning that it probably was not deposited slowly. "Good sorting is particularly significant because the sands are found in an environment where, unless deposition is very fast, one would expect silt and clay to be contributed..." 60
 Also, almost every sandstone layer exhibits some degree of sole
casts on its bottom surface as well as ripple marks on its top surface.
The upper surfaces of all of the sandstone layers, no matter how thick or thin,
were found to contain "asymmetric, linguoid ripples" According to Sheehan
"... these structures formed in response to unidirectional currents which
occurred either contemporaneously (at the same time) or penecontemporaneously
(immediately following) with sediment deposition." 60
Also, almost every sandstone layer exhibits some degree of sole
casts on its bottom surface as well as ripple marks on its top surface.
The upper surfaces of all of the sandstone layers, no matter how thick or thin,
were found to contain "asymmetric, linguoid ripples" According to Sheehan
"... these structures formed in response to unidirectional currents which
occurred either contemporaneously (at the same time) or penecontemporaneously
(immediately following) with sediment deposition." 60
Given all of these findings, what theory makes more sense? Were these layers deposited slowly where each layer was created over the course of tens, hundreds or even thousands of years, or were these layers formed rapidly by successive turbiditic flows in a highly silted watery environment? Is it reasonable for those such as Glenn Morton to suggest that burrowing creatures give evidence of a slow formation? What about the argument that such burrowing creatures must have been killed by each sandy turbidite so that a new colony of burrowing creatures would have had to take over the next layer of clay? This argument makes no sense at all. Since when does a few centimeters of sand kill any burrowing creature? This argument sounds almost silly, especially if one has ever tried to bury such creatures under sand at the beach. They simply dig out in short order. But, what about the fact that no evidence of "escape burrows" with "little circular piles of sand with a central crater pocking the entire upper surface of the sand" can be found? No one who considered that the tops of each sand layer shows current ripples would ask such a question because the watery current would surely have removed any such piles of sand in short order as soon as they were made. With each new sandy turbidite the burrowers would simply burrow up through the sand to populate the newly forming layer of organically rich material as it rapidly formed over the turbiditic sand flow in a heavily silted environment. More and more layers would have formed in rapid succession leaving no time for the bioturbation of lower layers. 60
We are left then with the curious findings of thin crisp alternating layers of shale and sandstone showing no evidence of bioturbation between layers and increasing layer thickness as one moves up these formations. This sort of layering is only consistent with rapid formation and cannot be explained by the prevailing paradigm where millions of years are required to produce such shale bed formations. (Back to Top)
 However, what about those ancient desert sand dune layers in the
Grand Canyon?
The popular science of today declares that the Coconino Sandstone layer
(third from the top) used to be an ancient desert formed over eons of time.
It is interesting that most of the layers in the Grand Canyon are felt to have
formed under water. However, the Coconino Sandstone layer is felt to be an
exception to this rule. This theory would mean that a layer of
water-deposited mud (the Hermit Shale) was followed by a layer of wind-deposited
desert sand over the course of between 5 and 10 million years, and then the area
was again covered by water and the Kaibab limestone was deposited.
However, what about those ancient desert sand dune layers in the
Grand Canyon?
The popular science of today declares that the Coconino Sandstone layer
(third from the top) used to be an ancient desert formed over eons of time.
It is interesting that most of the layers in the Grand Canyon are felt to have
formed under water. However, the Coconino Sandstone layer is felt to be an
exception to this rule. This theory would mean that a layer of
water-deposited mud (the Hermit Shale) was followed by a layer of wind-deposited
desert sand over the course of between 5 and 10 million years, and then the area
was again covered by water and the Kaibab limestone was deposited.
The Coconino Sandstone layer is quite interesting indeed.
It averages 96 meters in thickness (315ft.) and covers an area of 200,000
sq. miles to include most of northern Arizona from the Magollon Rim, northward
to the Utah border. It is up to 1,000 feet thick at its southern edge and
thins to a few feet at its northern boundary. The total volume of sand is
estimates to be approximately 10,000 cubic miles.62 The sand grains
themselves are fine grained, well rounded and sorted, and composed almost
entirely of quartz with no silt or mud contamination, just like most
The popularity of the desert origin for the Coconino Sandstone began with the work of McKee in the early 1930s.74 McKee initially focused on the physical qualities of the sandstone to support his conclusions that the dunes had been wind and not water deposited. Later he studied the footprints and concluded that they were most likely formed on dry sand.75,76 However, according to Leonard Brand, "Sedimentary features that were formerly thought to be diagnostic of eolian deposits are now known to be non-diagnostic. Stanley et al. (1971) pointed out that "grain frosting is no longer considered a criterion of wind transport," grain size distribution statistics have been ambiguous (for the Navajo), and "it can no longer be assumed a priori that large festoon cross strata prove an eolian dune origin for the Navajo or any similar sandstone because of the essential identity of form and scale of modern submarine dunes or sand waves, as documented during the past decade" (e.g., see d'Anglejan 1971; Harvey 1966; Jordan 1962; and Terwindt 1971)." 72
But what about the fact that the Coconino Sandstone
has preserved crisp footprints in delicate detail? Well,
does such detailed trackway preservation happen in dry desert sand?
When a lizard walks or runs over dry sand, what happens?
Footprint impressions are made, but nothing near the detail and crispness
that has been preserved in the Coconino Sandstone is produced.
Now, consider the likelihood that shifting sand will preserve very small
and delicate footprints made in dry sand. This seems
a bit hard to imagine. In fact, laboratory experiments in
real time have shown that the level of detail
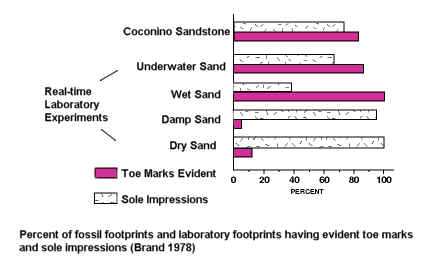 found in the Coconino Sandstone is best explained by the formation
of the tracts underwater or on sand that has been wetted and left standing
overnight. According to Brand's experiments the damp sand trackways that
did not stand overnight always had definite foot impressions but the toe marks
were rarely seen. The dampened surface formed a crust of sand that broke
apart into many small pieces when the animals walked over it. Sometimes
these pieces of crusted sand would be pushed up into a pile at the back of the
footprint or be scattered on around beside the footprint on the surface of the
sand. However, if the dampness of the surface of the sand was thick enough
so that it would not break up with the weight of the animal, the sand would
become rather hard and resistant to track formation. The only tracks
produced on this kind of wetted sand were a series of small dimples left by the
toes. The fossilized Coconino Sandstone tracks did not match the dry
or dampened sand tracks produced in the
found in the Coconino Sandstone is best explained by the formation
of the tracts underwater or on sand that has been wetted and left standing
overnight. According to Brand's experiments the damp sand trackways that
did not stand overnight always had definite foot impressions but the toe marks
were rarely seen. The dampened surface formed a crust of sand that broke
apart into many small pieces when the animals walked over it. Sometimes
these pieces of crusted sand would be pushed up into a pile at the back of the
footprint or be scattered on around beside the footprint on the surface of the
sand. However, if the dampness of the surface of the sand was thick enough
so that it would not break up with the weight of the animal, the sand would
become rather hard and resistant to track formation. The only tracks
produced on this kind of wetted sand were a series of small dimples left by the
toes. The fossilized Coconino Sandstone tracks did not match the dry
or dampened sand tracks produced in the  laboratory by Brand in several ways. The dry and damp sand tracks rarely
preserved toe marks or other details, while the fossilized tracks usually did
preserve toe marks. Dry sand tracks also had large ridges of sand behind
them which often flowed back into the previous footprint. Again, the
fossil footprints do not have these ridges nor were jumbled pieces of crusted
sand observed to be scattered around the fossilized tracks. The
proportions of the tracks were also different. Brand noted that the dry
sand tracks were longer than they were wide, but the fossilized tracks were
short in relative to their width. The tracks made in the damp sand were
simply too indistinct to allow adequate measurements.
laboratory by Brand in several ways. The dry and damp sand tracks rarely
preserved toe marks or other details, while the fossilized tracks usually did
preserve toe marks. Dry sand tracks also had large ridges of sand behind
them which often flowed back into the previous footprint. Again, the
fossil footprints do not have these ridges nor were jumbled pieces of crusted
sand observed to be scattered around the fossilized tracks. The
proportions of the tracks were also different. Brand noted that the dry
sand tracks were longer than they were wide, but the fossilized tracks were
short in relative to their width. The tracks made in the damp sand were
simply too indistinct to allow adequate measurements.
Brand also did experiments in which the slope of the sand rose above the water line. As the animals walked up out of the water their tracks changed as they went higher and higher from the waterline. Footprints close to the water level were poorly defined while those a little higher were crisp and clear as far as toe marks and sole impressions. However, as the animal progressed even higher to the more firm sand, the tracks became fainter and fainter until only toe mark dimples could be discerned. This transition effect from well-defined prints to vague toe marks or scratches is not seen in the Coconino Sandstone.
So why did Mckee believe that the tracks found in the Coconino Sandstone were made on dry desert sand? Mckee was heavily influenced in the formation of his desert theory by the influence of a paleontologist by the name of Peabody who told McKee that salamanders do not generally make tracks underwater but prefer to swim from place to place instead of walk. And, even when they do walk, they are partially buoyed up by the water so that their tracks are vague at best. McKee seemed to indicate that he had experimentally confirmed these suggestions made by Peabody. Thus, McKee (1947) concluded that the fossil tracks preserved in the Coconino Sandstone were most similar to the dry sand trackways produced in his own experiments because only in dry sand were any definite prints of individual feet formed. What is strange though is that there is no documentation of how extensive McKee's observations were as far as observing salamanders and their swimming or walking habits while under water.
Brand, on the other hand, documents that all five species in his study walked on the bottom sands underwater more than they swam from place to place through the water as long as they had a sandbar or some place in the testing tank where they could rest. Brand noted that this behavior is also observed in the field. When walking along the sand underwater all five species selected by Brand produced distinct footprints with toe marks and occasional sole impressions all along their trackways. Some of the prints also had ridges of sand pushed up behind them, but these ridges never extended back into the previous print. Brand concluded that, "The underwater tracks were most similar to the fossil tracks. Underwater trackways had toe marks as often as the fossil tracks, and they were uniform in appearance the full length of the sand slope, as the fossil tracks are. Also, the proportions of the fossil tracks were most similar to that of the underwater tracks." 72 However, Brand did leave open the possibility that such trackways could have been formed on a special type of wetted sand that had been wetted for several hours (overnight in his experiments). Based on this evidence many argue that the desert was wetted on occasion by light mists, dew, or a heavy fog. This allowed the various creatures living in this ancient desert to make their crisp trackways, which were subsequently covered by dry sand and preserved. Other dry-land features, such as raindrop impressions, crisp and steep leeward dune fracture faces and cracks in the sand, and the preservation of spider trackways are often cited as evidence in support of this dry-land formation hypothesis in opposition to Brand's underwater hypothesis. This dry land hypothesis quite reasonable in many respects that seem to require open air exposure, but there are still a few other very puzzling features that do not seem so consistent with a true desert-like environment or dune formation.
 What is rarely mentioned
in the literature is that the vast majority of the Coconino trackways all head
uphill.66 Evidently the
lizards/amphibians, arthropods, spiders and other creatures living in ancient
deserts did not like going downhill much at all. Also,
trackways often start and stop suddenly without evidence of sand-shift or
disturbance - like the creature suddenly vanished into thin air (or swam off in
the water).66,67,68
What is rarely mentioned
in the literature is that the vast majority of the Coconino trackways all head
uphill.66 Evidently the
lizards/amphibians, arthropods, spiders and other creatures living in ancient
deserts did not like going downhill much at all. Also,
trackways often start and stop suddenly without evidence of sand-shift or
disturbance - like the creature suddenly vanished into thin air (or swam off in
the water).66,67,68
McKee attempted to explain the relative absence of
downhill trackways by suggesting that the animals tended to "slide" downhill,
thus obliterating their own tracks in the sliding sand. One might wonder why the
animals would slide downhill when they were doing do fine going uphill without
the sliding problem. Those who have ever visited areas with desert sand
dunes will find that trackways on such sand dunes go every which way.
Also, one would expect that wetted sand would be much more cohesive than dry
sand and preserve tracks just as crisply no matter which direction the creatures
were heading. And, just in case there was any doubt, Brand performed a few
more experiments. Brand actually went to the trouble of inducing his
experimental animals to walk both downhill as well as uphill. On
underwater sand, wet sand, and damp sand, almost all downhill trails produced
easily recognizable trackways. On dry sand, the trackways of salamanders
were less well defined, but still relatively well 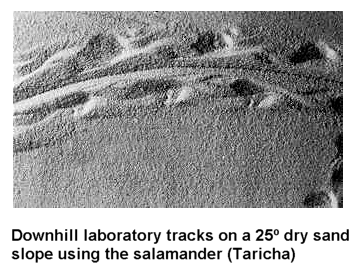 preserved, while that of lizards were still quite distinct (both walking or
running slowly). Only when running very fast did their tracks become
unrecognizable. Thus, the almost complete absence of downhill tracks in
the Coconino Sandstone layers seems to remain a mystery if they are truly desert
formations. It almost seems as though the creatures were trying to escape
something, like rise water levels, and that is why they were all generally going
uphill?
preserved, while that of lizards were still quite distinct (both walking or
running slowly). Only when running very fast did their tracks become
unrecognizable. Thus, the almost complete absence of downhill tracks in
the Coconino Sandstone layers seems to remain a mystery if they are truly desert
formations. It almost seems as though the creatures were trying to escape
something, like rise water levels, and that is why they were all generally going
uphill?
Brand also noted one other unusual aspect of some of the Coconino trackways. On occasion, there would be trackways that would head directly across a given slope at one angle or another, but the toe marks of both the back and front feet would be pointed up the slope. It seems unlikely that the animals that made these trackways would have walked sideways for such distances. Some have argued that desert lizards sometimes walk sideways in order to angle themselves to reduce the absorption of heat radiated from the scorching desert sand. The problem with this argument is that the sand was wet and therefore relatively cool - certainly not scorching hot. Others have suggested that a strong wind blew the animals sideways. This idea requires a very strong wind indeed to blow a relatively low profile lizard or salamander sideways in an even pattern for significant distances. Brand suggests that the more likely explanation is that these animals were walking in an underwater current, which seems at least plausible.
Also, the architecture of the Coconino sand dunes is not like that of modern sand dunes in modern deserts. The Coconino sand dunes have an average slope angle of 25 degrees while the average slope angle of modern desert dunes is 30-34 degrees (the resting angle of dry sand).69 Sand dunes formed by underwater currents do not have as high an average slope angle as desert dunes and do not have avalanche faces as commonly as deserts dunes do. Some crisp avalanche faces are found in the Coconino Sandstone dunes suggesting that at least some exposure to open air occurred, but such exposure may have been intermittent and relatively brief. Still what explanation can be given for the microscopic pitting and frosting of the grains of sand? It turns out that desert sand is not the only sand that can be pitted and frosted. The chemical process of sand cementation in the forming of sandstone can also cause pitting and frosting.70
So, it appears that the evidence does not fit the classic dry desert formation of the Coconino Sandstone layer over millions of years of time. Many of the trackways may even have been formed underwater or at best on long standing damped sand dunes where all the creatures walked only uphill. Ocean currents can and do make very pure quartz sand dunes with specific characteristics that match the dunes in the Coconino Sandstone.71 Heavy ocean currents can in fact amass huge quantities of sand in a very rapid timeframe. The sand dune angle found in the Coconino Sandstone layers would require a depth of water of around 300 feet and a fairly brisk current. In such a scenario, large dunes with cross bedding can be made very quickly.
 Consider also that there is no significant erosion between the
Coconino Sandstone layer and either the layer above it (the Toroweap Formation)
or the layer below it (the Hermit Formation). All of these
layers formed like sheets of glass - one on top of the other.77
Isn't it strange that significant portions of these layers have not been
weathered away to be filled in by overlying layers in an uneven way?
In fact, large contraction cracks penetrating deep into the Hermit Formation
(just below the Coconino layer) are filled in with Coconino sandstone. If
the Hermit Formation took millions of years to form, which would surely turn the
layers in this formation into solid rock in a small fraction of this time, how
did such deep cracks form in solid rock in such a way that the surface was
completely flat and yet the cracks themselves were filled with pure Coconino
sandstone? One would think that if such formations and characteristics
took long periods of time to form that the boundary between the Hermit Formation
and the Coconino sandstone would have been blurred by "bioturbation", disturbed
in an uneven way by erosion, and that the cracks found in the Hermit shale would
have been filled with other contaminants besides pure Coconino sandstone.
But, these findings are not strange if the layers were all formed
relatively rapidly by water deposition instead of over vast expanses of time.
Which theory has better explanatory value? (Back
to Top)
Consider also that there is no significant erosion between the
Coconino Sandstone layer and either the layer above it (the Toroweap Formation)
or the layer below it (the Hermit Formation). All of these
layers formed like sheets of glass - one on top of the other.77
Isn't it strange that significant portions of these layers have not been
weathered away to be filled in by overlying layers in an uneven way?
In fact, large contraction cracks penetrating deep into the Hermit Formation
(just below the Coconino layer) are filled in with Coconino sandstone. If
the Hermit Formation took millions of years to form, which would surely turn the
layers in this formation into solid rock in a small fraction of this time, how
did such deep cracks form in solid rock in such a way that the surface was
completely flat and yet the cracks themselves were filled with pure Coconino
sandstone? One would think that if such formations and characteristics
took long periods of time to form that the boundary between the Hermit Formation
and the Coconino sandstone would have been blurred by "bioturbation", disturbed
in an uneven way by erosion, and that the cracks found in the Hermit shale would
have been filled with other contaminants besides pure Coconino sandstone.
But, these findings are not strange if the layers were all formed
relatively rapidly by water deposition instead of over vast expanses of time.
Which theory has better explanatory value? (Back
to Top)
Subjective Taxonomic Classification
Since a single gene pool can produce "drastic" differences in phenotypic forms, how are scientists so sure of their fossil classification models? Often only slight phenotypic differences are enough to place a fossil creature in a different species, genus or even family group than its modern-day counterpart or than its counterpart found elsewhere in the geologic column. The problem is that differences, even fairly significant differences, are known to exist between members of the same gene pool. Because of this fact, taxonomic classification models can be quite subjective and even misleading.
For example, scientists from Berkeley have noted that, "the planktonic larvae of many marine invertebrates are commonly described as separate species when they are first discovered in the ocean. Only later when they can be reared in the laboratory can the link to their adult form be recognized. Similarly, the different life stages of many fungi are given different names because they have different physical forms and hosts. Only through detailed inoculation studies can mycologists work out which forms are members of the same life cycle. Since some fungi may have more than five discrete life cycle stages, this can be a long process. Similar problems exist for some marine algae and multiple-host parasitic organisms of many kinds. Even among well-studied vertebrates, some tropical birds have been described as separate species until they are observed to mate and rear young together."24
"Detailed study of large sympatric populations and fossil assemblages of the highly variable species Elphidium excavatum (Terquem) [Benthic foraminifera] collected from 20 widely spaced locations indicates that a variety of morphotypes of Elphidium can be linked to one another in a number of interlocking intergradational series. Ten morphotypes are recognized and grouped as formae (ecophenotypes) of Elphidium excavatum (Terquem); these morphotypes were previously considered as 22 independent taxa by various authors. Although all of these formae belong to the same species, it is suggested [by the authors] that the distinction among them should be retained because of their potential as a valuable interpretive tool in paleo-ecological and biostratigraphic studies of Holocene and Pleistocene sediments."25
"Because they are based on different stages in the life-cycle, fossil dinoflagellates and living dinoflagellates have largely received two sets of names, the equivalencies of which are becoming increasingly well known. For example, Gonyaulax spinifera and related species are known to produce cysts assignable to the genus Spiniferites. Indeed, it is generally informally acknowledged that Spiniferites and Gonyaulax are taxonomic synonyms. For several reasons this synonymy has not been formally proposed: 1) the fossil generic name Spiniferites is senior to the extant name Gonyaulax and acceptance of the synonymy would bring considerable changes to the nomenclature of this major extant genus (and conservation of Gonyaulax would cause a reciprocal chaos among fossil names); 2) the exact correspondence of Spiniferites species with Gonyaulax species is not clear; and 3) it is impossible to establish whether earlier representatives of the genus Spiniferites were cysts with a thecate stage identical to living Gonyaulax. In other words, to many researchers, it is useful and desirable to retain both Gonyaulax and Spiniferites while acknowledging that they may represent the same biological taxon."26
The naming of hominid fossils not immune from this subjective problem. In a March 2002 statement, Tim White, who co-directs the Laboratory for Human Evolutionary studies said, "There's been a recent tendency to give a different name to each of the fossils that comes out of the ground, and that has led to what we think is a very misleading portrayal of the biology of human evolution. But, when you find a fossil like this one so similar to Asian and European ones, it indicates the same species." "This whole species question is all about what you accept as a sharp enough distinction to tell you that it is a separate species," said Susan Anton, a Rutgers University anthropologist. "This particular skull is not going to solve that problem."27
Specific hominid fossils, such as the Solo fossils, have presented a bit of a problem as far as classification in concerned. "When they were first discovered, von Koenigswald believed them to be "tropical Neanderthalers." In 1963, Bernard Campbell classified them as Homo sapiens soloensis. Santa Luca, in 1980, classified them as Homo erectus erectus, with Milford Wolpoff declaring that they were not Homo erectus. Still others called them "archaic Homo sapiens." Because of their obvious similarity to the other Japanese and Chinese "classic" Homo erectus material, most investigators today recognize them as Homo erectus. The Solo fossils do, however, have a larger cranial capacity than does the average Homo erectus skull. For this reason, many evolutionists could not resist the temptation to consider the Solo people as "transitional" between Homo erectus and modern humans. Unfortunately, since evolutionists believe that modern humans arrived on the scene by 100,000 YBP, transitional fossils at 27,000 YBP will not fit. It is now known that there are many late-date Australian fossils almost identical to the Solo (Ngandong) people."28
The classifications of plants is classically prone to give different names to very similar plants or even parts of the same plant. Bill DiMichele, a paleobotanist, notes, "The problem of organ association is one of the reasons why paleobotanists insist on so many different names for isolated parts of the same whole plant. Furthermore, there are phenotypic convergences that can cause great confusion, such leaves of virtually identical morphology borne on ferns and seed plants. Separate names for each fossil plant organ can be carried to extremes, however, and not all paleobotanists, myself included, favor the attribution of separate names to organs otherwise known in attachment (yes, this is still done routinely, no kidding)."29
The Mazon Creek flora is incredibly diverse. Over 400 species from at least 130 genera have been identified from Mazon Creek nodules. However, the number of different kinds of plants represented is very difficult to determine. There are at least two reasons for this difficulty. The first reason is the convention among paleobotanists that separate plant parts receive different names. This procedure tends to inflate the number of plant names. The second reason is that paleobotanists are still trying to determine which taxa are valid.30
According to Meyen and Traverse the problems of naming fossils are as follows. "1. Living plants are assignable to a single taxon at any rank whereas fossil plants with dispersed parts and no observable original connections may be referred to several taxa of the same rank and have different names (Stigmaria, Lepidodendron, Lepidostrobus) 2. In living plants, all individuals belonging to a species belong to the same genus, etc. whereas in fossil plants various specimens of a species may or may not belong to the same genus and the genus may belong to different families when the complete plant is considered (Stigmaria may belong to genera assigned to Lepidodendraceae, Sigillariaceae, or Lepidocarpaceae). 3. Living plants are assigned to a complete hierarchy of taxa whereas fossil plants may be assigned only to genera with higher rankings unknown (some leaf genera might belong to pteridosperms, ferns, or cycads). 4. Living plants cannot be assigned to different genera based upon different types of preservation whereas fossil plants may be. 5. Different ontogenetic phases of the living plant do not normally serve as a distinction for a taxon whereas in fossil plants this is possible (seeds, microspores, megaspores, cysts). They concluded that fossil plant nomenclature requires only two special circumstances be reflected in the ICBN: 1) the possibility to keep genera of fossil plants outside the hierarchy of formally named higher taxa; and 2) the possibility to retain names of taxa established for various parts." 31,32
As it turns out, "Intraspecific variation is ubiquitous in systematic characters, yet systematists often do not deal with polymorphism explicity. For example, morphological systematists typically exclude characters in which any or "too much" polymorphism is observed, and molecular systematists often avoid intraspecific variation by sampling a single individual per species. Recent empirical studies have suggested that polymorphic characters contain significant phylogenetic information but are more homoplastic than fixed characters. Excluding polymorphic characters decreased accuracy under almost all conditions examined, even when only the more variable characters were excluded. Sampling a single individual per species also consistently decreased accuracy. Thus, two common approaches for dealing with intraspecific variation in morphological and molecular systematics can give relatively poor estimates of phylogeny."33 (Back to Top)
 Foraminiferans are actually
protozoans (single celled animals) of the Order Foraminiferida. Their
fossilized remains have been commonly used to relatively date sedimentary rock.
This practice is especially extensive in the oil industry. These creatures
are usually quite tiny, but have a fairly wide range of size from 0.1 millimeter
(mm) to almost 20 centimeters (cm) in length with an average that is less than 1
mm. They also have an extraordinarily fast doubling time of 3.65 days. The
parts of the creatures that are preserved in the fossil record are their shells.
Actually, these shells are not really shells in that they are
"intra-ectoplasmic" structures called "tests". These tests function
somewhat like a skeleton and are formed of small interconnected chambers.
The chamber connections are small openings between each chamber. The
openings are called "foramina" from which the name "foraminifers" comes from.
Foraminiferan shells or tests come in many shapes and sizes and have many
different designs to include simple tubes, straight series chambers, coils of
chamber and even complex labyrinths. Their walls can be formed by
particulate matter picked up from the surrounding environment or they can be
formed of pure calcareous material that is secreted by the foraminifer.
Foraminifers interact with their environment through pores and apertures that
exist in the test wall. Their taxonomy is based first on the wall
mineralogy and microstructure, then on chamber arrangement, apertural shape and
position, and finally ornamentation.
21,22
Foraminiferans are actually
protozoans (single celled animals) of the Order Foraminiferida. Their
fossilized remains have been commonly used to relatively date sedimentary rock.
This practice is especially extensive in the oil industry. These creatures
are usually quite tiny, but have a fairly wide range of size from 0.1 millimeter
(mm) to almost 20 centimeters (cm) in length with an average that is less than 1
mm. They also have an extraordinarily fast doubling time of 3.65 days. The
parts of the creatures that are preserved in the fossil record are their shells.
Actually, these shells are not really shells in that they are
"intra-ectoplasmic" structures called "tests". These tests function
somewhat like a skeleton and are formed of small interconnected chambers.
The chamber connections are small openings between each chamber. The
openings are called "foramina" from which the name "foraminifers" comes from.
Foraminiferan shells or tests come in many shapes and sizes and have many
different designs to include simple tubes, straight series chambers, coils of
chamber and even complex labyrinths. Their walls can be formed by
particulate matter picked up from the surrounding environment or they can be
formed of pure calcareous material that is secreted by the foraminifer.
Foraminifers interact with their environment through pores and apertures that
exist in the test wall. Their taxonomy is based first on the wall
mineralogy and microstructure, then on chamber arrangement, apertural shape and
position, and finally ornamentation.
21,22
Foraminifers are not only abundant in the fossil record but live today as well. They can be found practically anywhere where there is a body of water to include deep sea trenches, shallow seas and oceans, and even fresh water lakes. However, different types of foraminifers occupy different types of environments. For example, planktonic and pelagic species live in open water at various depths while benthic species live near, on, or in the ocean floor. In deep ocean environments, calcareous (made of calcium) material is dissolved, so foraminifers in this environment tend to be of the type that agglutinates surrounding material to make their tests instead of forming calcareous tests. Foraminifers move around via "pseudopodia" or hair-like extensions of protoplasm and have been clocked at several millimeters per hour. These are also used as "arms" for the gathering of food and building materials and for attachment to other objects. Sometimes the pseudopodia are even used to aid in floatation, as is the case with planktonic species. Foraminifers can also tolerate extreme environments to include very low oxygen levels, hyper- and hypo-salinity, as well as extremes of pH and temperature. What is interesting is that these different environments play a role on the type of framiniferan that occupies a particular area. For example, areas with low oxygen levels have foraminifers with a more flattened shape to increase absorption surface area. Their walls also tend to be thinner, more porous, and less ornamented. 22
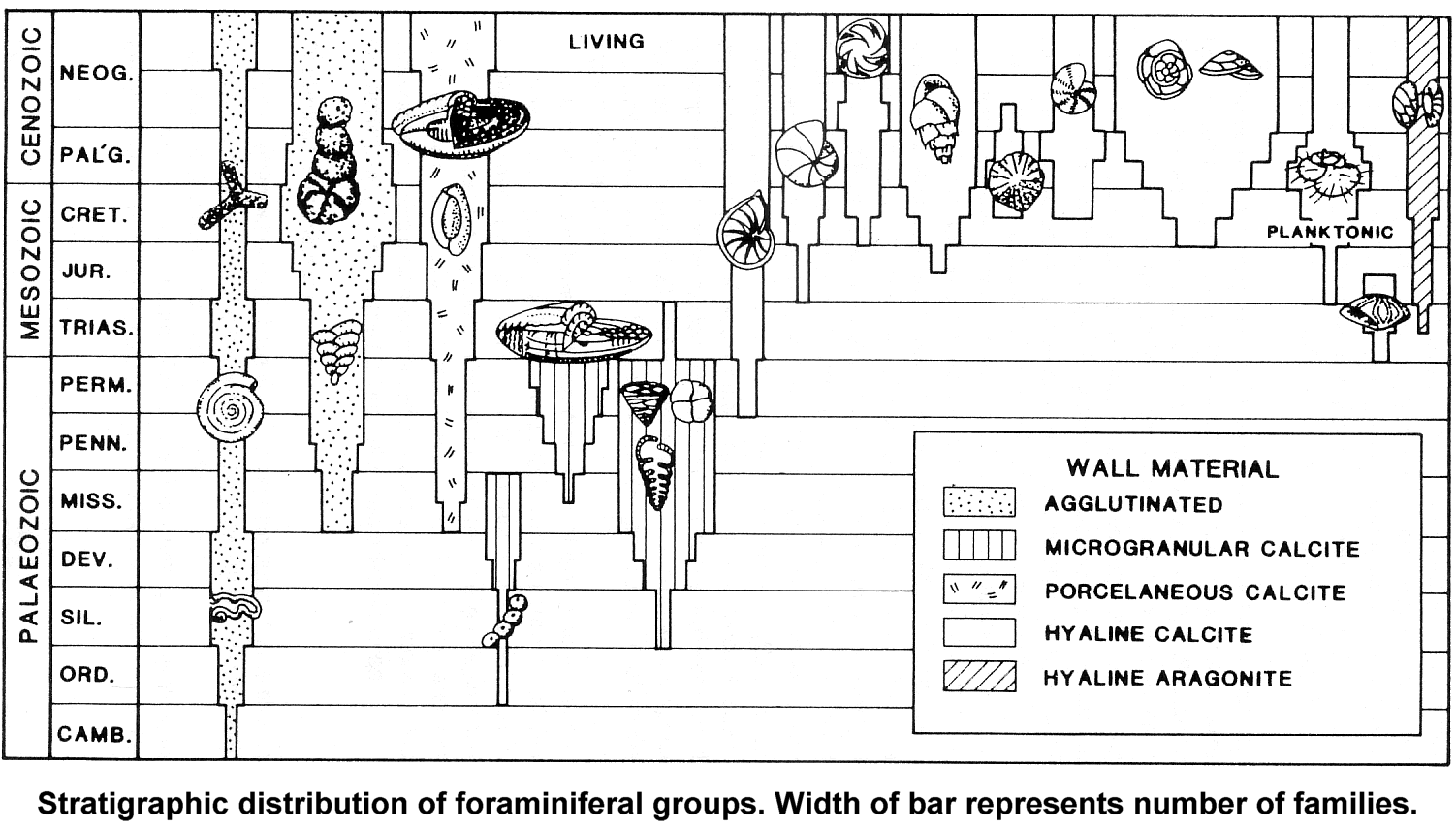
So, what do these very interesting creatures have to do with the theory of evolution? Well, as previous hinted at, different foraminifers are found in different layers within the geologic column. In fact, some are so closely associated with a given layer that when they are found, they practically define that layer within the geologic column. This association is so reliable that oil drilling companies rely heavily on the identification of these creatures as they drill their wells to the proper depth. Obviously then, if the layers in the geologic column represent long periods of historical time, the differences in foraminifers represent evolutionary changes over time. The layers could not possible have been formed rapidly by a watery catastrophe such as are mentioned in flood legends, for how would these foraminifers, who are similar in size and density, be sorted so neatly into the various layers in which they are found? There are some, such as the geologist Glenn Morton, who argue that:
"Foraminifera are small, single cellular animals which would have existed in the oceans prior to the flood and due to their small size should be found all mixed together in the same or closely related strata... Given the small size of the average species, they should all sort out at about the same time from the waters of the flood with the largest at the bottom and the smallest at the top. This is not what we find when we look at the foraminifera fossil record. Genera of forams, all possessing the similar shape and similar size and only differing in the details of the test decoration, are found over vast vertical distances in the geologic column... It is like throwing similar size and density sand particles, which are colored different colors, into a river and having the colors all sort out. This is impossible. Yet forams are so sorted. The only conclusion can be that their order is not due to a global flood but to a long period of deposition in which the animal life changed." 21
Morton's argument is especially interesting when one considers that different foraminiferan test decoration are found within the same species depending upon environmental factors of the local habitat. Tammy Tosk, also a geologist, argues that both habitat preference as well as environmental factors can have a rapid impact on foraminiferan morphology.22 Consider the following illustration and note that the foraminifers of today vary in morphology according to changes in ocean depth.
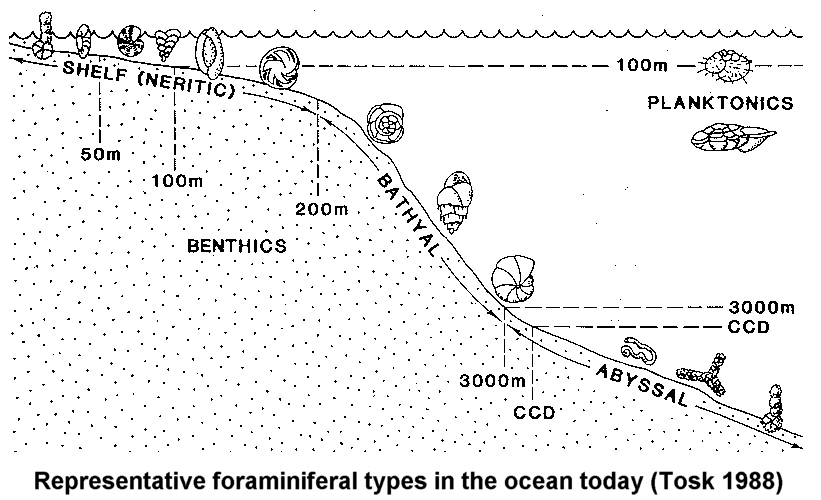
As illustrated above, Tosk argues that morphologic variation or "sorting" within the geologic column can be based on normal ecologic distribution. Tosk goes on to argue that within a single foraminifer species, certain members may have thickly ornamented tests under normal oxygen concentrations and thin less-ornamented tests in environments where oxygen concentrations are low. Such variations that are based, not in genetics, but in environmental influences, are called "ecophenotypic" variations. Based on these ideas, Tosk theorizes about how the geologic foraminiferan data could be explained by a rapid catastrophic burial:
"Because of the many examples of variation in living and fossil forms, foraminifers are considered to be extraordinarily plastic (Kennett 1976). A foraminifer may contain enough genetic information to express many different forms, depending on the conditions... A significant problem arises because similar forms are classified differently if they occur at different stratigraphic levels. These cases are explained as iterative evolution, that is, the same form evolved repeatedly through geologic history. Thus classification is subjectively influenced by evolutionary theory. Repeated occurrences could be explained as easily by a catastrophic flood model. If the foraminifers found fossilized at various levels in the geologic column were living at the same time in different ecologic zones, species common to several ecologic zones would be found at several levels. Gaps in the record only indicate that the species was not present in the source area or the ecologic zone being buried at that time, not that it was totally extinct. No coincidence of repeated extinction and identical evolution is required.
Ecologic zonation as developed by Clark (1946) would mean that foraminifers living in the lower seas or deeper parts of the ocean would be buried first as the sediments were redeposited by the gradually rising flood waters, while those from higher ecologic zones would be buried later The fossil record seems generally consistent with this model. [The figures above] show the distribution of foraminifers today and of fossils in the geologic column. Simple agglutinated forms that now live in environments ranging from the deep sea to estuaries, are found fossilized in Early Paleozoic and younger strata. Calcareous benthic species now predominate both in bathyal environments and in Mesozoic strata of the past, and presently floating planktonic forms from a higher ecologic zone are abundant in the higher Cenozoic strata of the past.
In the oceans today, calcareous material is dissolved below the carbonate compensation depth (CCD) usually at a depth of about 4000 m, depending on carbon dioxide concentration. Neither benthic nor planktonic calcareous foraminifers are generally found below that depth on the abyssal plains or in deep sea trenches, because their calcareous shells would be dissolved. Agglutinated forms are dominant
Agglutinated species are common in the Lower Paleozoic, and the benthic calcareous foraminifers found generally have thicker walls than forms higher in the geologic column. They could have lived near the pre-flood CCD where most calcareous forms, especially thinner-shelled planktonic species, would have been completely dissolved. Lower Paleozoic foraminifers are consistent, therefore, with the distribution expected by a catastrophic flood.
The fusulinids in the Upper Paleozoic, however, are an anomaly. Some species of fusulinids grew to volumes of more than 100 m3 (Ross 1979). Foraminifers which grew that large today have symbiotic photosynthetic algae living in their tests, and so must live within tens of meters of the ocean surface where sunlight is available. Large foraminifers from other groups live in shallow water tropical environments today; therefore, the fusulinids are interpreted also to have lived in a similar environment (Ross 1979), yet we do not find them at the top of the geologic column. Possibly they grew at the surface of pre-flood bodies of water of low altitude (Figure 1).
Planktonic foraminifers are not found in Paleozoic or Lower Mesozoic deposits. Even though living planktonic foraminifers float and would not be expected to be found in the early flood deposits, tests of those which had died before the flood should have been on the sea floor and should have been buried with those living there. Either they were not present in those ecologic zones, or they were not preserved as fossils. Because they have thinner, more porous tests than benthic forms, they could easily have been dissolved preferentially on the sea floor before the onset of catastrophic flooding, if their shells sank below the CCD.Benthic hyaline calcareous foraminifers become abundant in the Mesozoic. Triassic and Jurassic foraminifers are generally not as well preserved as later forms. In Cretaceous strata, both benthic and planktonic forms are diverse and abundant, making it correlative with the upper bathyal zone of the ocean today.
Foraminifers older than the Cretaceous are generally widely distributed. A Triassic species may be found in both Australia and Idaho, but nowhere in between (Tosk and Andersson 1988). Cretaceous and younger foraminifers have distribution patterns correlative with modern assemblages (Sliter 1972). Under the prevailing paradigm, this would mean that the pre-Cretaceous seas were more cosmopolitan because modern hydrographic patterns and ecologic distributions had not yet developed. Continental fragmentation and sea-floor spreading during the Cretaceous are used to account for the development of modern oceanic patterns at that time.
In a flood model, however, this pattern is what would be expected. During the more violent stages of the flood events, foraminifers from a small area would be scattered widely over the earth. As the violence of the flood died down, foraminifers would not be transported as far and might even begin developing their own ecologic distribution patterns. Major deposition during and after the Cretaceous could have become localized in basins and at continental margins. Life for foraminifers may have returned to normal in less affected areas." 22
 So, it seems at least possible to conclude that the apparent "sorting" of
foraminifers in the fossil record could have occurred outside of slow process of
millions of years involving gradual evolutionary changes. Morphologic
variations seen today are often based on ecologic environment that are fairly
distinctly "sorted." Arguments such as Morton presents need not
necessarily or even preferentially support the long age scenario over a
catastrophic formation of much of the geologic column.
So, it seems at least possible to conclude that the apparent "sorting" of
foraminifers in the fossil record could have occurred outside of slow process of
millions of years involving gradual evolutionary changes. Morphologic
variations seen today are often based on ecologic environment that are fairly
distinctly "sorted." Arguments such as Morton presents need not
necessarily or even preferentially support the long age scenario over a
catastrophic formation of much of the geologic column.
But what about arguments that Morton also raises that foraminiferans show smooth evolution between species? Consider the series of slowly changing forms presented at the right. Morton refers to this series as a clear example of plankton evolution in action by saying,
"One can clearly see the gradual transformation of the earliest into the latest species. This gradual change is imperceptible... Gradualistic evolution is documented among these tiny creatures laying bare the false claim that there are no transitional forms. What it shows is that the flood-advocates don't read anything except their own literature." 21
Well, after reading Morton's "literature" it seems that such variations in morphology might be easily explained by variations in environment. These forms pictures at the right might in fact be members of the same species of plankton. After all, foraminifer are quite "plastic" indeed. The genetic information contained in one common gene pool seems quite capable of producing startling morphologic variations. Considering this ecophenotypic variation potential, how then is Morton so sure that "gradualistic evolution" is taking place here? (Back to Top)
 Some point to the obvious vertical order of pollen found throughout the fossil
record and suggest that such sorting can only be explained by evolutionary
changes over time.21 It is true that pollens are also sorted in
the fossil record. However, to say that this sorting
is impossible for a large complex fluvial event or a relatively rapid series of
rather complex catastrophic events to explain, seems to be a bit presumptuous.
Some point to the obvious vertical order of pollen found throughout the fossil
record and suggest that such sorting can only be explained by evolutionary
changes over time.21 It is true that pollens are also sorted in
the fossil record. However, to say that this sorting
is impossible for a large complex fluvial event or a relatively rapid series of
rather complex catastrophic events to explain, seems to be a bit presumptuous.
As it turns out, different pollens can be water sorted based on various characteristic differences such as size, specific gravity, density, chemical composition (ie: hydrophobic vs. hydrophilic), and "fall velocity" in water. For example, most deciduous pollen types sink in water in a few minutes while bisaccate grains (pine pollen) float for much longer times, sometimes years, before they sink." Pohl (1933) noted that pine pollen could retain its buoyancy for up to four years.
Hopkins (1950) results show that there is some differential buoyancy between Pinus species. The grains that sank are those with smaller or deformed/broken air sacs. When placed on a water surface in a large water tank, over which a breeze of 13km/h was blown, conifer pollen drifts at a rate of 0.16 to 0.32 km/h. When oak pollen (Quercus palustris) was put through a similar process most of it sank within the first meter.
Bisccate grains will only sink if the bladders are pierced either by physical, chemical or microbial processes. They are therefore more likely to be saturated if they enter turbulent flow, or if they have been resident in a position (such as the soil surface) where they are open to microbial attack prior to introduction to the water.
Many pollen grains have mechanisms to prevent desiccation, but cannot prevent water uptake. Sporomorphs, many of which have no openings in the exine, have no inlet for water. Some, such as Lycopodium, also have an oily coat (Balick & Beitel, 1988). Attempts to saturate Lycopodium in the laboratory have shown that such grains are extremely resistant to water uptake. Extreme methods have been invoked, such as placing the sporomorphs in a beaker in a vacuum for up to eight hours (own method) or boiling in water (Reynolds 1979). This means that they are likely to remain afloat in the water, in nature, for long periods of time. This is consistent with results from the Volga River where Fedorova (1952) found Lycopodium spores over 7000km from their source.
Several authors have found that water currents can sort sporomorphs. This was first seen in the field by Muller (1959) who was looking at the palynomorph distribution in recent sediments in the Orinoco delta area. He noted that the pollen of Rhizophora, which is relatively small, was carried further out to sea than larger sporomorphs from similar source areas.
Cross et al. (1966) also found evidence of sorting in a marine setting as lighter, more buoyant, grains were carried further toward the southern end of the Gulf of California. Davis et al. (1971) and Davis and Brubaker (1973) explained differences in the distributions of oak (Quercus) and ragweed (Ambrosia) in the sediments of several small North American lakes as resulting from sorting. The smaller ragweed grains remain in suspension for longer periods of time, allowing them to be transported to the littoral areas of the lake by wind generated currents. Sorting was also used to explain results presented by Chen (1987) from the sediments of lake Barine in Queensland, Australia. Larger grains are deposited in the littoral areas, while smaller grains are carried to the lake center by these currents.
Clearly sorting of sporomorphs can occur in many depositional environments. The behavior of sporomorphs as sedimentary particles is, however, poorly understood. In a more recent study of alluvial sediments from a canyon stream in the chuska mountains of Arizona, Fall (1987) tested the assumption that the pollen in the alluvium was from local vegetation. She found that pollen concentrations in general were highest in the fine-grained sediments. Non-arboreal sporomorph types however were found in higher numbers in the sandy sediments. Similar results have been reported from Glacio-lacustrine sediments in Alaska (Goodwin, 1988) and braided stream deposits adjacent to the Caribou River in Yukon (Catto, 1985) or they may be due to different source areas of tributaries involved. These results suggest that the deposition of fine-grained sediments, sporomophs in particular, is related both to the flow conditions of the water and the bed material over which the water flows.
From the evidence presented here is it suggested that, where water currents drop below 35cm/sec, sporomorphs may be sorted. This sorting is primarily due to size, but sporomorph density is also important." 23
Pollen, Spores and Vascular Plants in the Cambrian and Pre-Cambrian
It is often argued that the pollen or spores of modern plants are limited to the modern or Tertiary (often Eocene) layers of the geologic column. Despite such common assertions, there seem to be at least a few long-standing problems with this notion. Although the subject of the Salt Range beds is proscribed among Indian and many western paleontologists today, the case rests precisely where it did in the early 1950s (Ghosh et al. 1951). The fossils are modern in aspect ("Eocene" according to Sahni 1944) yet the beds containing the fossils are overlain conformably by early Cambrian sediments (Coates et al. 1945).
In the mid- 1940s there was a lively controversy about the age of the saline series of the Punjab salt range. On the basis of field evidence, geologists like Gee and Fox regarded it as Cambrian and Pre-Cambrian. But on the basis of the microfossils comprising Shreds of gymnospermous and angiospermous woods, cuticle of grasses, etc., obtained from Saline Series by macerating, Prof. B. Sahni regarded it as Eocene. His contension was that angiospermous remains could not have occurred earlier than the Tertiary. To strengthen his argument he cited the absence of vascular elements in the Cambrian strata of the Salt Range which he said were unconformably lying over the Saline Series. Prof. Ghosh and his associates checked the results obtained by Prof. Sahni and his collaborators and found the occurrence of vascular elements even in the Cambrian rocks.
The controversy over the age of the saline Series could not be resolved due to the sudden demise of Prof. Sahni in1949. But it inspired Prof. Ghosh to further investigate whether vascular plants existed in Cambrian times or not. To collect sufficient evidences, he and A. Bose, his main associate in this study, investigated Cambrian rocks of Punjab Salt Range and Kashmir, Pre-Cambrian rocks of Dharwar and Cuddapah, some Vindhyan rocks and a few Cambrian specimens from North America. From these rocks they obtained wood elements and monolete and trilete types of spores, leading to the conclusion of the fact that their findings were not in agreement with the view of the majority of investigators who did not consider vascular plants to have existed in Pre-Silurian times. Still they had the courage to stand by their own observations, which were also supported by the works of Naumova, Reissinger and Kopeliovitch in Europe and Jacob and his associates in India. In 1969 they again published a paper on the spores of vascular plants obtained from nine samples of Cambrain rocks of North America, further reiterating the existence of vascular plants in the Cambrian.
Prof. Ghosh's investigations related with the occurrence of earliest vascular plant led him to have a more critical looks to at all the organic remains reported from Pre-Cambrian deposits. This formed the subject matter of the 43rd Acharya Jagadis Chandra Bose endowment lecture which he delivered in November 1973 under the title "Life in the Proterozoic". . . The controversy received global attention and Ghosh became known as "Cambrian Ghosh" worldwide . . .
On his retirement from ONGC in 1962, Ghosh became an emeritus professor in the Botany Department of Calcutta University where he advised several palynology students, e.g., Manju Sharma, A. Nandi. 84, 85-96
Also, in 1956 S. Leclercq published a paper in the journal Evolution entitled, "Evidence of vascular plants in the Cambrian" that included the following introduction:
While the information given by palynology agrees with the flora known in the Upper and Middle Devonian, the assemblage of spores found in the Lower Devonian, Silurian and Cambrian plead in favor of a more varied and advanced type of flora than the plant impressions have so far suggested. The recent discovery of Lycopodiaceous shoots in the Middle Cambrian of East-Siberia appears to confirm the evidence of palynology investigation. 97
Some researchers have reported finding pollen of higher plants in strata shown by standard dating methods to be extremely old. These findings call into question the whole conventional account of the evolution of plants. In one instance, parties of scientists in Venezuela reported finding pollen of flowering plants in Precambrian rock formations judged to be 1.7-2.0 billion years old. This posed a serious problem, because according to current theory the flowering plants evolved fairly recently, only 100 million years ago. The story is as follows:
Late in 1963 U. C. K. Dunsterville made an expedition to collect orchids around Cerro Venamo, at the westernmost point on the frontier between Venezuela and British Guiana (where this mountain is known as Wenamu Head). He noted some shale‑like beds at the base of a towering cliff of Roraima sandstone and collected samples for their possible paleontological interest. G. Fournier, palynologist of the Mene Grande Oil Company, processed the samples and recovered well-preserved pollen and spores. Subsequently, L. Nijssen and J. A. Sulek, palynologists of Compa-Shell de Venezuela and Creole Petroleum Corporation, respectively, processed other pieces and recovered identical plant microfossils.
This discovery of pollen and spores in a formation of supposed Precambrian age was so remarkable that a reconnaissance expedition of qualified geologists was organized to verify the facts of the case. During April, 1964, with the assistance of personnel and vehicles of the Ministerio de Minas e Hidrocarburos, the locality was visited by a party which included N. Benain, P. J. Bermdez, A. Espejo, U. Fournier, A. Mendez, J. A. Sulek and F. Wright. They confirmed the salient facts as recorded by Dunsterville. The shale-like beds, being less competent, had eroded away below the massive Roraima sandstone, leaving an undercut extending 10-12 ft. inwards at the base of the cliff. The original samples were loose, weathered fragments from the talus slope below. New samples of unweathered rock were collected from the face of the undercut.
On their return to Caracas, the three palynologists made independent investigations of the new samples. Utmost care was taken to avoid any possibility of superficial contamination. The rock cleaves along finely laminated bedding planes which are coated with limonite. Every effort was made to avoid these planes and some of the pieces processed were the central nubs left after chipping away the external parts of large blocks of the rock, which was dense enough to sound when struck with a hammer. Nevertheless, microfossils of the same type as before were recovered. . .
Interpretation
As to interpretation of the significance of the fossil pollen and spores, two sharply divided opinions have been expressed. The writers make no attempt to adjudicate, but state the two concepts impartially.
One group adopts the attitude that the radiometric dating of dolerites and a hornfels within the Roraima Formation as Precambrian is beyond dispute, hence the pollen (and spores) must have entered as secondary contamination. The improbability that pollen could withstand the baking process, which converted shale to hornfels, is adduced as further evidence that the pollen must be allochthonous. The absence of macroscopic plant remains in the Roraima Formation is also noted, despite its assumed continental (fluviatile) origin. It is admitted that entry of the pollen into its present site defies simple explanation, though some form of washing in by meteoric waters in the geological past via joints in the overlying sandstone seems the most probable cause.
The second group holds that by no conceivable physical means could the pollen (and spores) have entered the metamorphosed sediments from the outside. They are dense impermeable rocks compressed by an overburden of hundreds of feet of the overlying Roraima sandstones. The undercutting at Cerro Venamo suggests that the cliff has been steadily retreating, hence the face which was sampled must have been deep within the formation until quite recent times. The Roraima sandstones are quartzitic, of low permeability, hence carriage of extraneous pollen through them by percolating water seems highly improbable. Even if this process could occur, entry of such pollen and spores into the nonporous hornfels lacks an explanation. Furthermore, if plausibility of this process be granted, it would have been operative for a long period, and a mixed suite of spores and pollen should be expected.
In counter-argument against the first group, it is claimed that the assertion that pollen and spores cannot withstand anaerobic baking of their parent shales has never been tested experimentally. As regards the radiometric dating, there is a disquieting overlap between stated ages of the Roraima Formation and the underlying basement rocks. The latter suffered complex deformation and vulcanism, and were then deeply peneplaned before being covered by thousands of feet of Roraima sandstones, and only after these prolonged events were the dolerites intruded, on which age-determination of the Roraima has been based. If the radiometric technique is valid there should be a long and clear-cut time-gap between ages assigned to the basement rocks and to the Roraima beds. Such a gap does not exist in the experimental results published, but this discrepancy is glossed over in the latest summary of radiometric dating in British Guiana.
As stated, we offer no solution to the paradox. It is clear, however, that botanist Dunsterville in his hunt for rare orchids stumbled on a highly intriguing geological problem. 98
So, to resolve the difficulty, one group of scientists decided that although the dates of the rock were correct the pollen must have been a recent intrusion, even though entry of the pollen into those layers defies simple explanation. The second group held that the pollen had been there since the rock had formed, but concluded that the dating was wrong and the rock was of recent origin. The two groups thus contradicted each other in their interpretations of the evidence. The real significance of this treatment is that both groups felt compelled to look for ways to avoid contradicting the standard story of evolution, to which they were strongly committed. An summary of this problem is as follows:
In the mid- 1940s there was a lively controversy about the age of the saline series of the Punjab salt range. On the basis of field evidence, geologists like Gee and Fox regarded it as Cambrian and Pre-Cambrian. But on the basis of the microfossils comprising Shreds of gymnospermous and angiospermous woods, cuticle of grasses, etc., obtained from Saline Series by macerating, Prof. B. Sahni regarded it as Eocene. His contention was that angiosperm remains could not have occurred earlier than the Tertiary. To strengthen his argument he cited the absence of vascular elements in the Cambrian strata of the Salt Range which he said were unconformably lying over the Saline Series. Prof. Ghosh and his associates checked the results obtained by Prof. Sahni and his collaborators and found the occurrence of vascular elements even in the Cambrian rocks.
The controversy over the age of the saline Series could not be resolved due to the sudden demise of Prof. Sahni in1949. But it inspired Prof. Ghosh to further investigate whether vascular plants existed in Cambrian times or not. To collect sufficient evidences, he and A. Bose, his main associate in this study, investigated Cambrian rocks of Punjab Salt Range and Kashmir, Pre-Cambrian rocks of Dharwar and Cuddapah, some Vindhyan rocks and a few Cambrian specimens from North America. From these rocks they obtained wood elements and monolete and trilete types of spores, leading to the conclusion of the fact that their findings were not in agreement with the view of the majority of investigators who did not consider vascular plants to have existed in Pre-Silurian times. Still they had the courage to stand by their own observations, which were also supported by the works of Naumova, Reissinger and Kopeliovitch in Europe and Jacob and his associates in India. In 1969 they again published a paper on the spores of vascular plants obtained from nine samples of Cambrian rocks of North America, further reiterating the existence of vascular plants in the Cambrian.
Prof. Ghosh's investigations related with the occurrence of earliest vascular plant led him to have a more critical looks to at all the organic remains reported from Pre-Cambrian deposits. This formed the subject matter of the 43rd Acharya Jagadis Chandra Bose endowment lecture which he delivered in November 1973 under the title "Life in the Proterozoic".
The presence of fragmentary vascular plant microfossils of the Vindhyans and their correlation with Salt Range could never be reconciled. Thus, Ghosh came in direct controversy with Birbal Sahni whose viewpoints contradicted Ghosh's. The controversy received global attention and Ghosh became known as "Cambrian Ghosh" worldwide. (Link & Link).
By 2001 at least, this problem had not been resolved. Here is an interesting presentation given by Michael Cremo in 2001 at the XXI International Congress of History of Science, Mexico City, entitled, "Paleobotanical Anomalies Bearing on the Age of the Salt Range Formation of Pakistan: A Historical Survey of an Unresolved Scientific Controversy."
"The age of the Salt Range Formation in the Salt Range Mountains of Pakistan was a matter of extreme controversy among geologists from the middle nineteenth century to the middle twentieth century. Of great importance in the later discussions were fragments of advanced plants and insects discovered in the Salt Range Formation by researchers such as B. Sahni. According to Sahni, these finds indicated an Eocene age for the Salt Range Formation. But geological evidence cited by others was opposed to this conclusion, supporting instead a Cambrian age for the Salt Range formation. Modern geological opinion is unanimous that the Salt Range Formation is Cambrian. But Sahni's evidence for advanced plant and insect remains in the Salt Range Formation is not easily dismissed. It would appear that there is still a contradiction between the geological and paleontological evidence, just as there was during the time of active controversy. During the time of active controversy, E. R. Gee suggested that the conflict might be resolved by positing the existence of an advanced flora and fauna in the Cambrian. This idea was summarily dismissed at the time, but, although it challenges accepted ideas about the evolution of life on earth, it appears to provide the best fit with the different lines of evidence. The existence of advanced plant and animal life during the Cambrian is consistent with accounts found in the Puranic literature of India." 108

Many will argue that
in different places throughout the word, there can be found layers of forests
one on top of each other, with their trees, "in the position of growth" (still
standing up).
It is also said that these forests each have their own layer of soil.
So, it is felt that each of these were forests grew over long periods of time on
top of previous forests, each of which was buried by some long ago catastrophe.
What is interesting about these places (ie: Yellowstone National Park
were up to 65 different layers can be found with trees in the vertical
position)13 is that the trees are still oriented in their
Another strange finding of the organic
layers is the fact that they are sorted in various ways. Take for instance
the fact that pine needles and leaves are not mixed together, but are found in
separate organic layers despite the fact that there are both pine and hardwood
trees "growing" from the same organic layer(s). Also, the are
proportionately less pine needles than there are leaves even in areas that are
dominated by conifers. In a real forest, conifers drop a much higher mass
of needles than deciduous trees drop leaves. And yet, in the fossil
forests of Yellowstone, needles are relatively sparse even at the bases of large
fossilized conifers. This interesting fact has been recognized as far back
as 1899 when Knowlton remarked about the absence of needles in the organic
levels associated with the large fenced petrified tree near Roosevelt Lodge in
Yellowstone National Park. 15
One would expect to find great numbers of sequoia needles and some cones, since
most of the upright trees are sequoia (70%). However, large numbers of broad
leaves and only a few pine needles are seen in the organic levels. Sequoia
needles were rare or absent. Although petrified sycamore stumps are not common,
leaves of sycamore are the most abundant broad-leaf fossils.
Pollen is another
problem. Trees with wind-transported pollen, such as walnut and sycamore,
should have left a pollen record in the forest floor, but little or no pollen of
these two has been found. Modern forest floors contain pollen in abundance
inversely proportional to the distance from the source trees especially trees
for which wind is the pollen-transporting agent. Research done on four levels of
Specimen Creek Petrified Forest showed no positive correlation between fossil
pollen abundance and the proximity of possible source trees.
The organic layers themselves also show no
significant decay as one goes from top to bottom. In a real forest floor,
organic material decays. Evidently, this does not happen in the forests of
the past because the leaves at the bottom of organic layers are just as well
formed as preserved as the leaves at the tops of these organic layers.
Also, the material between the leaves is sorted, course to fine as one moves
upward (Consider the figure at the right). Evidence for such water sorting
occurs in about 70% of the organic layers.
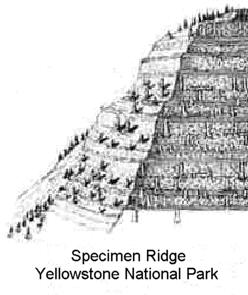 positions with each
other.
Their soil is also found to be water sorted (course to fine), and often
is found half way up a tree instead of at its base. This organic material also averages only 3 cm in thickness
and, for many of the "forest" layers it is missing altogether. In some
areas, such as Mt. Hornaday, as many as 43% of the forest layers have no organic
layer at all. The lower layers of Specimen Creek generally do have organic
layers (96%), but the upper layers of Specimen Creek have far fewer organic
layers. It turns out that the average "forest" without an organic layer to
is about 24%. It seems rather strange for a forest to grow into full bloom
without forming an organic layer. How is this explained?
positions with each
other.
Their soil is also found to be water sorted (course to fine), and often
is found half way up a tree instead of at its base. This organic material also averages only 3 cm in thickness
and, for many of the "forest" layers it is missing altogether. In some
areas, such as Mt. Hornaday, as many as 43% of the forest layers have no organic
layer at all. The lower layers of Specimen Creek generally do have organic
layers (96%), but the upper layers of Specimen Creek have far fewer organic
layers. It turns out that the average "forest" without an organic layer to
is about 24%. It seems rather strange for a forest to grow into full bloom
without forming an organic layer. How is this explained?

There is yet another problem with the intermittent burial of forests by volcanic activity and mud-slides. If a volcanic mud slide buried only the lower parts of the trunks of the trees of a growing forest (as might be suggested by the fact that some of the upright trees penetrate overlying layers), the taxonomic composition of the new forest that grew on this new surface would be similar to the composition of the forest that was buried. The cones, seeds, nuts, and fruits would fall from the unburied branches and foliage and repopulate the new surface with a similar forest. Such correlation between adjacent levels of the Specimen Creek Petrified Forest has not been found.18
The normal maturation of soil also involves the slow breakdown of feldspar and other minerals into clay. Out of 65 layers of Specimen Creek in Yellowstone National Park, only nine bands of clay were found among only seven organic layers. Horizontal sampling of two of the clay bands at 2.5 - 3 m intervals for 30 m showed a constant mineral distribution. Abundant unweathered feldspar is scattered throughout the Yellowstone organic levels, suggesting repeated rapid burials.19 None of the 58 organic levels outside the 9 bands of clay contained detectable amounts of clay. The apparent absence of clay in the majority of levels (implying that normal weathering of soil did not occur) raises questions about the passage of long time intervals between levels. This datum also questions the validity of the assumption that the organic levels (upon which trees with hundreds of rings sit) represent true soils. Furthermore, the sudden abundant appearance of clay in a few horizontal bands that included both organic levels and layers of clay in the associated breccia beds between levels suggests transport rather than in situ formation of the clay minerals.
Despite much study of the Yellowstone Petrified Forests, no animal fossils have been found. Why are animal remains absent from the plant fossil-bearing levels of Yellowstone? Because forests would be expected to harbor a wide variety of animals, some of which would be buried by the successive mud slides, the absence of animal fossils has been a mystery. Volcanic activity could have caused larger forest animals to flee elsewhere, but flight cannot be used as an explanation for the absence of all animal remains because many animals could not or would not leave their forest habitats. Land snails, some amphibians and reptiles, many insects, arachnids, and worms would not escape burial. Immature members of many types would be unable to flee. In addition, bones, eggs, teeth, scales, molted skins, castings, droppings, burrows, etc., would qualify as evidence of animal life. None of these have been found in the fossil forest organic levels during a century of research. Considering that delicate plant parts are excellently preserved, animal remains should also have been preserved if they were present. Only one exception is known. Remains of termites have been found in chambers within the petrified wood.19 If the petrified trees are standing where they originally grew and if the organic levels are the growing surfaces still intact and undisturbed, the absence of animal fossils is difficult to explain. If, however, the trees and the organic debris making up the soil levels were transported by water, the separation of animals from the plants before burial is much easier to explain.
The volcanic ash found in the various layers
is itself is quite interesting. The chemicals that make up the ash are
uniform throughout all the layers. In fact, there are only four unique
chemical patterns to be found in the ash of Specimen Creek. Spark source
mass spectrometry analysis of trace elements in the bands of ash revealed pulses
of ash from four source areas for the Specimen Creek Petrified Forest. The
four trace element profiles interweave in an irregular manner up the sequence of
65 organic levels of Specimen Creek Petrified Forest. If these 65 ash layers
(organic levels) were laid down over a long time span, the ash that was laid
down thousands of years later near the end of the series of ash eruptions would
have changed sufficiently to produce a new and different trace element profile
(a new "fingerprint"). This has not been the case. The rapid burial of the whole
sequence seems to be required. 
The root systems of the upright stumps seems
to be a main argument in favor of their
in situ growth. Of course, some of the petrified trees have broken
roots; but when were they broken? Even if a permit to collect petrified wood
within the park is obtained, excavation of stumps is not permitted; furthermore,
digging is difficult in the hard rock. Consequently, to determine if the root
breakage seen is pre- or post-petrification is difficult. Several examples of
abrupt root terminations from Mt. Hornaday, Mt. Norris, Tom Minor Basin, and
Specimen Creek strongly indicate that, at least in some cases, the tree roots
were broken before the trees were buried by volcanic gravels and muds. This
evidence supports the view that the trees were transported. However, small
rootlets can be located at the bases of upright stumps, and this feature has
been
If a forest were killed by a mud flow that buried the bases of the
trees, the tops of the trees would extend above the new ground surface. They
would overlap a new second forest that would commence growing on the new
surface. During the time of the growth of the new second forest (before it in
turn was buried by another volcanic mud slide) the old first forest snags would
have time to rot, to be infested with insects, and to break drown. Even the tops
of stumps that reached only to the root area of the second forest level (no
actual overlap) would also be expected to experience decay. The soil in which
the roots of the second forest grew would not be a good preserving medium for
the tops of the stumps extending up from below. One of the striking features of
the Yellowstone petrified trees is their good preservation. If pieces of the
petrified wood are prepared as microslides, the wood tissue may look nearly as
fresh as tissue from a living tree. Seldom do they exhibit any evidence of decay
and weathering.19
This suggests that the trees have not been subjected to these processes during
the passage of time.
The parallel orientation of the horizontal logs, mentioned earlier,
is better explained by water or mud transport. The dip of the beds from which
the trees arise seldom exceeds 7 degrees, which is not enough to cause all the
trees to fall downhill.19 Prevailing winds or volcanic blasts could
align fallen trees, but they would not cause the long axes of the cross-section
of the upright stumps to have a similar compass alignment. The asymmetry of the
cross-section of a stump, especially at its base, is usually a result of the
influence of major roots that cause flare to extend for some distance up the
stump. Volcanic lahars (fast-moving volcanic mud slides) or currents of water or
mud could be the forces that acted on roots and trunks to produce similar
alignment for both stumps and logs. Modern forests do not show the degree of
orientation that the fossil forests of Yellowstone show. This orientation
seems to speak for catastrophic transport and against in situ forest
growth.
One of the first observations made when
research commenced on the petrified forests was the barkless condition of both
the horizontal and upright trees. Subsequent examination has revealed some thin
layers of bark remaining on a few of the trees. In addition to the trees being
mostly barkless, all the branches have been broken off. Even large branches, 25
cm or more in diameter, have been removed. Only scoured stubs remain on the tree
trunks. Trees buried and later excavated by water erosion during the eruption of
Mount St. Helens did not have all the bark or limbs removed. Trees floating for
a period of time in turbulent water would more likely lose bark and branches due
to softening of the bark and abrasion.14,15 The eruption of Mount St. Helens and the
resultant flooding in locals such as Spirit Lake might offer some insights into
how the stacked forests of Yellowstone might have formed. The floating log
raft in Spirit Lake does in fact provided some insights on taxonomic sorting.
Most of the stumps sitting erect on the bottom of Spirit Lake are Silver Fir,
Nolle Fir, and Hemlock. Douglas Fir, abundant in the surrounding forests, has
only 2.2% representation. Sampling of the trees in the floating raft solved this
discrepancy; 48% are Douglas Fir. Apparently this species is more buoyant and
resistant to water saturation. Cedar was represented by 2.2% of logs floating in
the lake; yet the surrounding forests contained a higher percentage of cedars.
Sampling of the broken wood pieces along a transect on the shore gave 11% for
cedar. Some of the erect trees floating in the lake or standing on the lake
bottom are over 20 m tall. The argument that tall petrified stumps must be in
position of growth does not apply to a flotation scenario.
Some have argued that Spirit Lake is not a
good analog for the Yellowstone Fossil forests, because the large number of logs
accumulating on the bottom of Spirit Lake very different from the more scattered
fossil logs and stumps in Yellowstone. However, one important difference between
the two deposits is that Spirit Lake has not had adequate sediment input to bury
the sinking logs and stumps. If the 1980 Mount St. Helens eruption had been
followed by a series of volcanic breccia flows into Spirit Lake, spaced long
enough apart to bury successive sets of logs and stumps as they sank, it would
likely have produced a deposit very similar to the Yellowstone Fossil Forests.19 If this is not yet convincing, these same trees were
studied to see if they might have matching patters of growth in a tree-ring
analysis.
Matching patterns of growth would be highly unlikely if the trees lived
thousands of years apart in time. In 1991, Dr.
Michael Arct presented such a study for his PhD dissertation (building upon
research that he had been doing since 1979).
He sampled fourteen fossil trees at different levels in a twenty-three
foot section of the Yellowstone formations.
Analysis showed that all fourteen trees matched and that ten of them died
at the same time. The other four trees died seven,
four, three, and two years before the other ten died. The theories of ancient formation no longer seem to match the
facts available.7, 8
The Rest of the Story
Of course, although it took some time, secular scientists
have also begun to realize that the old notion of forests growing in place on
top of each other over vast periods of time is simply wrong. In fact, as far
back as 1985, secular geologist William J. Fritz compared Mt. St. Helens with
Specimen Ridge. In his book Roadside Geology of
the Yellowstone Country, which was available
in park bookstores, he shared a bit of excitement in the midst of otherwise dry
geological accounts: “When I visited the Mount St. Helens area shortly after the
eruption, it was just like Yellowstone!”. Both the mudflows and the appearance
of the trees, he added, looked identical.
Coppedge bought that book in a 1994 return visit, but
noticed that the new Hamilton’s Guide (1994) still repeated the old story. Aware
that the old sign at Specimen Ridge was gone, he asked a park ranger about it.
The ranger said that park geologists no longer believed that interpretation;
they were leaning to the new idea that mudflows carried the logs into their
current positions.
Coppedge took this photo of a fresh, new-looking sign at the Petrified Tree
exhibit near Tower Junction. Current sign at Yellowstone’s Petrified Tree
exhibit (9/06/15) Four things are notable about the new sign. The fossil forests are explained as the result of
catastrophic mudflows from massive landslides after volcanic eruptions. The sign admits that “Anatomically, the trunk is
indistinguishable from present-day redwoods in California.” The background picture shows similar trees from Mt.
St. Helens as an analogue to Yellowstone’s fossil forests. There is no mention of the old paradigm.
The closest
thing to an admission the old story was wrong is a National
Park Service page about fossils. There’s no
date on the page, but the latest reference is from 2013, so it had to be written
or revised after that. There’s an indication in the page source code that it was
last updated in January 2015. Around 1900, F. H.
Knowlton identified 147 species of fossil plants from Yellowstone, 81 of them
new to science. He also proposed the theory that the petrified trees on the
northwest end of Specimen Ridge were forests petrified in place.
Additional studies and
observations informed a modification of Knowlton’s original hypothesis.
Andesitic volcanic eruptions such as the 1980 eruption of Mount St.
Helens showed that trees are uprooted by rapidly flowing volcanic debris flows.
The volcanic debris flows not only transported the trees to lower elevations,
the trees were also deposited upright. Thus, with increased scientific
knowledge, our understanding of Yellowstone’s fossil forests has changed.
What this implies is that
Knowlton’s theory did not constitute “understanding.” It was a
misunderstanding—one that the park fed to the public uncritically for decades on
their interpretive signs.
It is quite interesting to note that fossilized trees are
not like one would expect from the normal evolutionary picture of what
happened.
According to current popular scientific belief many fossilized trees were
buried naturally either as they lay fallen on the ground or as they stood
growing. However, what is unique about almost all
fossilized trees is that they do not have roots or branches and little if any
bark.1, 3 Also, the ones that have fallen
and that are laying horizontally, align themselves in the same direction.1,
3 Also, many fossilized seashells and
tree leaves are oriented relative to each other throughout the fossil
record. The trees in the forests of today do not do
orient themselves when they fall.
When they fall, they fall in a fairly random way without a statistical
significance in their orientation. This is not so with
petrified trees. Petrified trees all line up.
Even the vertical ones have a particular orientation.1
What could have caused these trees to loose all their roots, branches, and bark
and to line up in the same direction as every other tree in that layer?
During the floods of Mt. St. Helen's eruptions, the trees did exactly the
same thing. Such "current" orientation is found throughout the fossil
record, with orientation of fossils being observed over vast areas, sometimes
hundreds of thousands of square miles. Although not all fossils show
current orientation, a sizable percentage do show such current orientation.
Clearly, such orientation on such massive scales as are found throughout the
fossil record are not consistent with slow sedimentation and burial, but rapid
burial by the heavily silted currents of a large and sometimes massive fluvial
event or series of closely spaced events. (Back
to Top)
Powder River coal seams run remarkably thick and unsullied
by other material. Usually, unwanted sediment such as clay washes over a
deposit before coal seams can get very thick. But Power River coal is packed in
immense strips, some more than 200 feet thick. These seams stretch vast
distances up and down the basin. "They're hundreds of miles long. They're fifty
miles wide." says James McClurg, a geologist at the University of Wyoming.
"They're not little pods of an acre or two. They're immense things."
McClurg, who has studied the basin for more than a decade, says "No other place
in the world has as many seams 50 feet or more thick. But, the Powder
River basin is not only an economic resource. To geologists, it's also an
intriguing scientific enigma. Geologists have been studying the
basin for more than a century, largely to answer a baffling question: How did
the seams get so massive? Or more precisely, why weren't the seams diluted by
influxes of clay and other impurities before they thickened?" He adds, "It
would be like blindfolding yourself, spinning around, and hitting the center of
a dartboard one-hundred times in a row."5 (Click on Video Below)
This is quite a mystery considering the fact that a seam
of coal 200 feet thick would have taken a layer of organic material over 1,200
feet thick to create it. That is a lot of peat to be
in just one area. But, what makes this even more
unbelievable is that the Powder River Basin covers an area of over 10,000 square
miles. This problem could be solved with the idea of
a massive flood deposition of huge quantities of organic material in a very
rapid timeframe.
Similar deposits of huge amounts of plant material occurred during the
Mt. St. Helen's eruption and flooding by Spirit Lake. (Back to Top)
Dinosaur
Tracks In Coal
The current prevailing theory is that these dinosaurs
walked around in a swamp and their footprints left impressions in the peat.
These prints were then filled in by sand and clay and preserved.
It does not seem likely that soft surface peat would hold a print long
enough to get filled up.
Even so, if this is actually what happened, then should not the print
look more like a solid cast than an imprint?
In other words, if a print were filled in with sand or clay and thus
preserved, should there not then be a solid form of that mold?
It seems rather like the clay layer was formed first and then the prints
were made as the animals walked over it. Also,
the tracks do not seem to be all throughout the peat as it might be expected if
the popular theory were correct, but only in those layers where the vegetation
appears to have been covered rapidly by repeated waves of inorganic
sediments.
The tracks were then made through this layer of sediment and pressed into
peat itself to be preserved as coal.4 Consider also that the coal itself is
very pure throughout. There simply are not enough impurities in the coal
to support the notion that the layers of vegetation were formed slowly over a
long period of time. Again, this is interesting in the light that the
footprints are only preserved in those layers of coal that have a thin overlying
layer of clay (Click on Video Below).
Starving
Dinosaurs Consider, for example, the
Morrison Formation
(Late Jurassic) in the western United States. This formation has an average
thickness of 100m (300 ft) and extends well over 1,000,000 square km (about
700,000 square miles), being found from Canada to Texas, the Dakotas to Idaho
and Arizona to Oklahoma. It is known as one of the
world's richest sources of dinosaur fossils, but also contains fossil fish,
frogs, salamanders, lizards, crocodiles, pterosaurs, dinosaur eggs, and shrew-
to rat-sized mammals.
The dinosaur bones occur in the middle green siltstone beds and in the
lower sandstones of the Morrison Formation, often in graveyards composed of
densely packed bones. It is commonly said that this
vast formation was created by sediments carried by streams and rivers from some
ancient highlands into swampy lowland environments.
But how did gentle streams and small intermittent floods pack together so
many dinosaur fossils as well as a wide variety of other fossils, without
preserving very many plant fossils as well?
It seems like the Morrison Formation is, "practically barren of plant
fossils throughout most of its sequence." Some have admitted that the, "absence
of evidence for abundant plant life in the form of coal beds and organic rich
clays in much of the Morrison is puzzling." 102
Another interesting feature of the
Morrison Formation, especially in locations such as Dinosaur National Monument
(DNM), are a group of fresh-water clams of the genus Unio. Some of
these clams are found in the articulated form with the two matching shells
closed and intact. This feature indicates that the cause of death for
these clams was rapid burial and represent a "transported death assemblage".103
The same sedimentary layers found in other locations, like Grand Junction,
Colorado, display a large number of these unionids, all articulated, and
generally recognized as having been "buried alive during an episode of rapid
sedimentation".104
The remains of whole dinosaurs,
unionid clams, snails, logs, and wood fragments from the Quarry sandstone, many
with stream orientation, all testify to some degree of watery transport. The
more easily transported bones like ribs and phalanges are under-represented in
DNM compared to less-easily transported items like femurs, which suggests the
winnowing action of water. Also, at the DNM quarry, the bones
are found in three distinct intervals within the 50-foot-thick, channel-shaped
Quarry sandstone. The three sandstone "channels" scour into the surfaces
beneath, and experts have struggled to imagine the kind of "rivers" that each of
the channels represent. The notion taught for decades at the Quarry Visitor
Center by DNM rangers, that dinosaurs were washed up on a point bar along the
bank of a meandering river, is now discredited.105 Bones are
especially concentrated in the bottoms, not the sides, of the scour channels.
The sand grains and pebbles in the sandstone are dominantly composed not of
quartz, the typical river sediment, but of altered tuff and chert fragments of
probable volcanic origin. The lowest of the three levels, where dinosaur bones
are most abundant, contains isolated larger pebbles dispersed in a sandy matrix,
a texture unlike that of normal rivers. The texture and composition of the lower
interval suggests deposition from a muddy suspension, not normal bedload
transport in a river. Mudflows associated with catastrophic floods during the
recent eruptions at Mount St. Helens volcano produced fluidized sediment
slurries in wide river valleys and deposited similar textures.106 The
upper two intervals of the Quarry sandstone, where dinosaur bones are less
abundant, have noteworthy scour surfaces with cross beds of sand and pebbles
indicating eastward transport of muddy and sandy sediment over large dune
structures by very fast water currents. We can imagine dinosaur carcasses
suspended buoyantly in a denser-than-water flow. How far they floated is
unknown, but the process of suspension may have not been very abrasive. Clams,
snails and logs were also moved with the volcanic pebbles and carcasses within
the slurry. As deposition of sediment and carcasses occurred, the remaining flow
became enriched in water going from a muddy, slurry suspension current to a
less-muddy traction current. The deposit itself gives the impression of
catastrophic watery deposition events - i.e., a massive flood.107 Also, fossils within this region
show general current orientation (Confirmed by work of Arthur Chadwick, Ph.D.).
It seems like such massive and concentrated burial grounds as are found
in the Morrison Formation that show current orientation with little associated
vegetation are best explained by very large catastrophic flooding events with
massive sorting and transport of large dinosaur bones and other fossil
assemblages.34,35 (Back
to Top)
Termite
Nests
Within the Jurassic Morrison Formation, thought
to be some 40 million years old, are interesting formation that have been
interpreted by mainstream scientists, like Steven Hasiotis, as "termite nests" (link).
Concerning the formations illustrated in the photo above, Hasiotis writes,
"Enormous pillars are interpreted as subterranean termite nests in alluvial and
eolian deposits. . . [These nests are] composed of thousands of simple and
compound galleries, 0.15 to 0.5 cm in diameter to pan-sized chambers,
preferentially preserved because of the organics used in construction."100
Some of these "nests" are truly enormous - - up to 40 meters in vertical size.
But, are they really termite nests?
The photo above is a close up photo of the wall of a real
termite next. Notice how the termites used small globs of mud mixed with
fragments of rock as well as fragments of vegetable matter, saliva, and feces
(even termite body fragments) to build their nests. These globs of mud are
then lined up with each other to form little hills and furrows in the walls of
the rooms and passage ways in the termite nest. As it turns out, the
formations claimed by those like Hasiotis to be termite nests do not have these
particular features they do not have evidence for termite walls made out of
little globs of mud in linear rows nor do they have the fossilized remains of
any sticks, twigs, leaves, fungi, or any other organic building material.
Rather, these fossil formations are comprised of purified quartz crystals which
is very similar in chemical composition to the surrounding "country rock" - as
per the work of Ariel Roth. 101 Under microscopic analysis, to
include scanning electron microscopic analysis, the crystals in the "tubes" in
the formations appear to gradually "merge" with the surrounding rock - rock
which also shows vague linear outlines with the same basic structure of the
"tubes" in the fossil "nests". Surrounding Country Rock Calcium Carbonate Concretions
Additionally, in the same region as the formations in
question, are the above pictured formations made out of calcium carbonate.
These inorganic calcium carbonate concretions also have very similar morphologic
features to the most striking features of the supposed termite nests which were
interpreted as tunnels and passage ways. See the following close up
pictures of these calcium carbonate concretions below to see if you can tell the
difference between them and the supposed "termite nests" found nearby.
Taken together, these interesting concretions found in the
Jurassic Morrison Formation seem more consistent with a non-organic process of
formation than with termite nests or any other process of organic formation or
true trace fossils of any living thing.
I will leave you with one last observation.
There is a common saying among scientist that "finding the head of a dinosaur is
like finding a needle in a haystack." 2
This is to say that the greatest majority of dinosaur skeletons found
have no head attached to their bodies. There are
actually only a dozen or so complete dinosaur skeletons in the entire world.
Many more dinosaur skeletons than this have been found of course, but
they are not complete. Often they are missing their
heads. Why would a dinosaur loose its head if it
died a simple death and just fell over on the ground to be buried?
Even scientists admit openly that it appears like most dinosaurs were washed
into their current locations by heavy currents.2
A head is not as well attached to the rest of the body as other limbs.
A catastrophic flood could be the reason that there are so many headless
dinosaurs around. (Back to
Top)
Dinosaur Soft
Tissues and Blood
This account was given by Mary Schweitzer, a PhD student at the
time, from Montana State University. A well preserved Tyrannosaurus rex
skeleton had been found in 1990 and brought for analysis to Montana State
University. During microscopic examination of the fossilized remains, it
was noted that some portions of the long bones had not mineralized, but were in
fact original bone. Upon closer examination it was noted that within the
vascular system of this bone were what appeared to be red blood cells (note
retained nucleus in the center of the apparent RBCs and the fact that reptiles
and bird generally retain the RBC nucleus while mammals, like humans, do not).
50 Of course, this did not seem possible since the survival of intact red
blood cells for some 65-million years seems very unlikely if not downright
impossible. Further testing of these cells was done to attempt to disprove the
notion that they could possibly be red blood cells. Several analytical
techniques were used to characterize the material to include nuclear magnetic
resonance (NMR), Raman resonance and Raman spectroscopy (RR) and electron spin
resonance (ESR). These techniques did identify the presence of heme
group
molecules ranging in size from between 5,000 and 30,000 daltons (between 35 to
over 200 amino acids in size), but the detection limits of these methods were
not able to rule-out or rule-in the presence of hemoglobin or myoglobin
proteins due to the small amount of specimen available. So,
Schweitzer and her team decided to use a more sensitive detection method to
detect certain very specific types of proteins. They used the immune system of
rats. They injected some of the T. rex extract into laboratory rats
to see if these rats would mount an immune response to the foreign T. rex
material. And, the rats did mount a very specific immune response against
hemoglobin. This immune response was not just against hemoglobin in
general, but against certain types of hemoglobin.42 The reaction was
strongest against pigeon and rabbit hemoglobin with a weak reaction against
turkey hemoglobin, but there was no reaction against snake hemoglobin. The
specificity of these reactions were further confirmed by the lack of reactivity
with plant and sandstone extracts. This is significant since many previous real time studies with
protein decay suggest that sizable portions of protein sequences do not remain
intact beyond a few tens of thousands of years at best (Fossils, Protein and
DNA). Proteins and DNA usually decay very rapidly. Therefore, it would be
quite something indeed to find significantly intact hemoglobin after tens of
millions of years. Some have even suggested that significantly intact proteins
would indicate that such fossils are not in fact tens of millions of years old,
but were in fact recently buried within only a few tens of thousands of years at
most. Consider the conclusions that Schweitzer and her team made
concerning these findings:
"The production of antibodies specific for hemoglobin in two
rats injected with the trabecular extract is striking evidence for the presence
of hemoglobin-derived peptides in the bone extract. . . That the antisera did
not react with snake hemoglobin shows that the reactivity is specific and not
artifact. . . When considered as a whole, the results support the hypothesis
that heme prosthetic groups and hemoglobin fragments were preserved in the
tissues of the Late Cretaceous dinosaur skeleton." 42
Schweitzer goes on to suggest that "Immunogenicity is not dependent
on fully intact protein, and even very small peptides are immunogenic when
complexed with larger organic molecules . . . even after extensive degradation
has occurred."42
But how extensively, roughly, could the hemoglobin molecules have degraded and
yet retain their ability to elicit a fairly strong and quite specific immune
reaction in laboratory rats? In order to obtain such strongly specific
immunogenicity it would seem that a significant percentage of the globin
portion of the hemoglobin molecule would need to be intact in at least
some of the extracted specimen. In fact, larger molecules were indeed present.
Schweitzer proved that there were heme-containing molecules ranging from at
least 5,000 to 30,000da in size, and maybe even larger. But, how could a protein
of any significant size large enough to elicit an immune response to begin with,
as well as a specific immune response observed in this case, be maintained over
the course of 65 million years? One might very reasonably conclude that
natural decay, over this amount of time, would completely destroy the ability of
hemoglobin or the required larger fragments of degraded hemoglobin from being
antigenic much less so specifically antigenic. The explanation for this phenomenon, given later by Dr. Horner
(Schweitzer's boss) and even Schweitzer herself, was that the tougher heme
molecule survived the 65 million years with maybe three or four amino acids of
the original globin molecules attached to it. Consider the following
statement Schweitzer made in a response to an inquiry by Dr. Jack Debaun:
"But the heme itself is too small to be immunogenic [only about
652 daltons]. We believe that there were possibly 3-4 amino acids from the
original protein attached to the heme, and that was what may have spiked the
immune response." 43
There are several reasons why I feel
this way. For one thing, a certain
minimum antigen size is required before it can elicit an immune response
regardless of its structure. The most potent immunogens are macromolecular
proteins with molecular weights greater than 100,000da (~740aa - Note: the
average amino acid weighs ~135da).
Substances weighing less than 10,000da (~75aa) are only weakly
immunogenic, and those foreign proteins/antigens weighing less than 1,000da
(~7aa) are usually completely non-immunogenic. Homopolymers (repeats of the same
amino acid) are pretty much non-immunogenic regardless of size.
Co-polymers of glutamic acid and lysine must be ~35,000da (~250aa) to be
immunogenic. It seems then that, in general,
immunogenicity increases with structural complexity.
Also, aromatic amino acids, such as tyrosine or phenylalanine, contribute
much more to immunogenicity than do non-aromatic amino acids.
For example, the addition of tyrosine to a co-polymer made up of
glutamate and lysine reduces the size limitation to ~15,000da (~100aa) and
adding tyrosine and phenylalanine together reduces the minimum to 4,000da
(~30aa). Also, it is all four levels of protein
structure (1o, 2o, 3o, & 4o) that
influence immunogenicity - not just a short linear sequence of amino acids.44-47 Of course, a rather specific immune response can be elicited by
relatively few amino acids as part of an epitope on a larger protein molecule,
but they usually are not immunogenic without first being part of a larger
molecule (i.e., "complexed" to a larger molecule before being introduced to the
immune competent host). Also, antigenic epitopes are
not usually sequential in nature but are assembled by protein folding (i.e.,
bringing together amino acids that are widely separated on the protein chain).
This means that a rather large portion of the original molecule usually needs to
be intact in order for most epitopes to remain intact.
Epitopes with definite three-dimensional shapes and charged amino acids are
particularly well recognized by antibodies. The average epitope probably
involves about 7 to 15 contact amino acid residues and a few of these may be
critical to the epitope's specificity and the avidity of the antibody-antigen
reaction.44-47 But, in order to make an epitope antigenic, it
must be processed first. Antigen presenting cells (APCs) like macrophages, dendritic cells,
and even B-cells are responsible for antigen processing and the presentation of
epitopes/antigens to the T-cells. T-cells do not
recognize the initial foreign antigen directly.
They only recognize processed parts of antigens, usually consisting of no
more than 15 or so amino acids, presented to them by APCs in association with
MHC (major histocompatibility) molecules. So, in
order to activate T-cells (required for cellular immunity and very helpful in
humoral immunity), the foreign antigen must first be recognized as "foreign" by
the APC cells. This initial APC recognition requires
more than just a handful of amino acids floating around or else there would be
complete meltdown of the immune system.
In fact, generally speaking, molecules with a molecular weight less than
10,000da (~75aa) are only weakly immunogenic when picked up by APC cells.
Significant potency usually requires antigens to be rather
large at over 100,000da (~750aa), or at least above 5,000da (~35aa).48,49 Given all this, it seems quite difficult for me to imagine how "3
or 4" amino acids stuck to a heme group could elicit an immune response in the
first place, not to mention a specific immune response.
Recall that the heme molecule, by itself, only has a molecular weight of
around 652da. To make a strong as well as specific
immunogen (such as the immunity developed in rats exposed to T. rex
extract in this case) one might expect the immunogenic hemoglobin molecules to
be at least 5,000 to 10,000da (~35 to 75aa or so) in size.44-47
Certainly then, a heme group with 3
or 4 amino acids attached to it (just over 1,000da) would not seem to give rise
to such an immune response (specific to a certain type of hemoglobin) observed
by Schweitzer et al. in rats exposed to T. rex
bony extract. There are several common arguments used to
try and explain these apparent difficulties. One argument is that very small
degraded fragments of hemoglobin molecules (3 or 4 amino acids in size)
reattached themselves to each other to form new larger molecules with sufficient
size to be immunogenic. The problem here is that the rearrangement of so many
covalent bonds over the course of some 65 million years would seem to affect not
only the inter-amino acid bonds, but the intra-amino acid bonds as well. Of
course, such a degree of rearrangement would destroy antigenic epitope
specificity. Note also that the heme molecule itself is not bound by covalent
bonds to the globin portion of the hemoglobin molecule. It is held in place in a
crevice of the globin molecule by many non-covalent bonds that are individually
much weaker than the covalent bonds that hold the amino acids together. If the
covalent bonds were so commonly broken over time so as to leave only very small
chains of 3 or 4 amino acids from the original globin molecule intact, how was
the much weaker non-covalent bond between the heme group and a small chain of
only 3 or 4 amino acids maintained? It would seem that as the protecting crevice
surrounding the heme group was degraded to any significant degree that the
non-covalent bonds holding the heme group molecule in place would not have
lasted very long. The heme group would have been lost long before the much
stronger covalent bonds between amino acids in chains much longer than 3 or 4
amino acids would have been broken. Another argument is sometimes used that
Schweitzer failed to identify any specific size of hemoglobin fragment by gel
electrophoresis. What happened is that the electrophoretic pattern
observed by Schweitzer was a diffuse or smeared pattern. This means that
there were no discrete clusters of proteins that were the same size.
Certainly, this is only to be expected since a wide range of protein sizes would
be formed after an extended period of degradation. The fact of the
matter is that hemoglobin fragments ranging between 30 and 200 amino acids in
size where definitely present in the T. rex extract (per NMR analysis
filtering) and that these molecules were most likely intact fragments of the
original T. rex hemoglobin molecules and not reformed molecules made up
of tiny fragments.42 This find calls into question not only the nature of the
fossilization process, but also the age of these fossils. How such soft tissue
preservation and detail could be realized after 68 million years is more than
miraculous - - It is unbelievable! Schweitzer herself comments that, "We may not
really know as much about how fossils are preserved as we think." 54
Now, if that is not an understatement I'm not sure what is. So,
it seems rather clear, despite the objections of many
evolutionists, to include Schweitzer herself, that a 1,000da molecule would
elicit an extremely weak response at best and would not necessarily elicit a
specific response to a certain type of hemoglobin molecule since surface
epitopes are generally more specific in their antigenic nature than are buried
epitopes (i.e., heme is somewhat hidden within a cleft of the hemoglobin
molecule so 3 or 4 amino acids attached to it would also be somewhat hidden). How then is it remotely logical to suggest that a molecule weighing
just over 1,000da (a heme group plus 3 or 4 amino acids) could elicit such a
strong as well as specific immune response as Schweitzer et al. observed? In light of the additional recent finds of even more striking
soft tissue and blood cell preservation, it
seems much more likely that such an immune response so specific for certain
types of hemoglobin could only be elicited by a larger portion of intact
hemoglobin than many scientists seem to even consider. Of course, one
can't really blame them because explaining how delicate soft tissue vessels
(with obvious red blood cells inside containing relatively large portions of
hemoglobin molecules) could remain intact for over 65 million years seems just a
little bit difficult. All of this is a rather mute point, of course, in
light of the fact that T. rex
collagen has been subsequently sequenced.
The new finding will be viewed
skeptically, admitted one of the researchers involved in the two studies. "It's
very, very, very controversial because most people have gone on record saying
there's an absolute time limit to anything that's protein or DNA," said Mary
Schweitzer, a molecular paleontologist at North Carolina State University. Matthew Carrano, a dinosaur
curator at the Smithsonian Institution
in Washington, D.C., who was not involved in either study, said the protein
findings are robust. "Here are the pieces of the protein. If you're going to
refute this you have to explain how these pieces got in there," Carrano said in
a telephone interview. "It's not another molecule mimicking the protein and
giving off a similar signal. This is the actual sequence." 99 Of course, despite this surprising turn of events,
scientists do not question the notion that the dinosaur bones really are tens of
millions of years old. They still assume the long ages and evolutionary
relationship that they assumed before - as per the following passages:
A comparison by Asara's team
of the amino-acid sequence from the T. rex collagen to a database of
existing sequences from modern species showed it shared a remarkable similarity
to that of chickens. Amino acids are the molecular building blocks of proteins;
there are 20 of them used by organisms to build proteins, and their precise
order is determined by instructions found in DNA. "I'm grateful that he was able
to get the [amino acid] sequences out. That's the Holy Grail," Schweitzer told
LiveScience. . . Until now, family trees have been constructed from the
shapes of bones and teeth, a not-always-reliable technique." This finding supports the idea
that chickens and T. rex share an evolutionary link and bolsters previous
research showing that birds evolved from dinosaurs and that birds are living
dinosaurs. "Here we have a real molecule
from a real dinosaur, and it's much more similar to a bird than it is to
anything else," Carrano said. 99
Such finds are much more consistent with a fairly recent
catastrophic burial within just a few thousand years of time. Non-catastrophic
burial would allow for rapid biodegradation of such delicate soft tissues. Time
itself destroys soft tissues as well as DNA and proteins in short order.
Current real-time observations suggest that bio-proteins could not remain intact
more than a few tens of thousands of years - 100,000 years at the very outside limit of protein decay. The fact that
such proteins are found, intact, in bones supposedly older than 65 million years
is simply inconsistent with such an assumed age - by a few orders of magnitude.
Mary
Schweitzer's 60 Minutes Interview
Conclusion So, where does the evidence put us?
What position seems most reasonable? What theory
answers the most questions? Do fossils and the
geologic column that contains them represent millions of years of slow
sedimentation or do they reveal a time of huge catastrophe and rapid deposition,
death, and burial on a global scale? (Back to Top) Veith, W. J., Amazing Discoveries Video Series, 2000. (
http://www.amazingdiscoveries.org
) National Geographic On Assignment, Lucy the
T-rex,
2000. Miller, Alisa C., Fossilized Trees of
Petrified Forest National Park, from the
website: www.geo.arizona.edu, Nov. 22, 1998. Walter Brown, Robert Gentry, Mike Beesley, David Gentry,
Lonnie Melanshenko,.The Young Age of the Earth,
Video Series, 1993. Link1 (about two
thirds of the way through the video and merging into the next video)
Link2 (The first
few minutes of the video) Earth Magazine, Power River Coal: Geologic
Enigma, Environmental Dilemma, May 1993. Veith, W. J., The Genesis Conflict, 1997. John Morris, The Young Earth, Master
Books, Colorado Springs, CO, 1994, pp. 112-117. Michael J. Arct, Dendroecology in the fossil
forests of the Specimen Creek area, Yellowstone National Park, Ph.D.
Dissertation, Loma Linda University, 1991; Dissertation
Abstracts International 53-06B:2759, 1987-1991. Lemonick, M. Gene Mapper, Time, Vol.
156, No. 26, pp110, 2001 M. Schweitzer and T. Staedter, 'The
Real Jurassic Park', Earth , June 1997 pp. 55-57. Morell, V., Dino DNA: The hunt and
the hype, Science 261(5118):160-162, 9 July 1993. Schweitzer Mary, Proceedings of
the National Academy of Sciences, Vol. 94, p 6291. National Park Service. 1980.
Petrified forests of Yellowstone. US Department of the Interior, Washington, DC.
Handbook 108, p 6-9 Coffin HG. Orientation of trees
in the Yellowstone Petrified Forests. Journal of Paleontology 50:539-543, 1976. Coffin HG. 1979. The organic levels
of the Yellowstone Petrified Forests. Origins 6(2):71-82 Coffin HG. 1983. Erect floating
stumps in Spirit Lake, Washington. Geology 11:198-199. Coffin HG. 1983. Origin by Design.
Washington DC:Review and Herald Publishing Assn., p 143. DeBord PL. 1979. Palynology of the
Gallatin Mountain "fossil forest" of Yellowstone National Park, Montana:
preliminary report. First Conference on Scientific Research in the National
Parks. US Department of the Interior, National Park Service Transactions
Proceedings Series 5:159-164 Coffin, Harold G. 1997.
TheYellowstone Petrified "Forests". Origins 24:5-44. Coffin HG. 1987. Sonar and scuba survey of a submerged
allochthonous "forest" in Spirit Lake, Washington. Palaois 2:179-180. Morton, Glenn. 2000. http://www.glenn.morton.btinternet.co.uk/micro.htm Tosk, Tammy. 1988. Origins 15(1):8-18. Holmes,P.L. (1994): The sorting of spores and pollen by
water: experimental and field evidence. - In: Traverse,A. [ed.]: Sedimentation
of organic particles, 9-32, 10 figs., 1 tab.; Cambridge: Univ. Press. Greuter, W. et al., 1996. Taxon 45: 349-372.
http://www.versiontech.com/origins/news/news_article.asp?news_id=113
http://www.ngdc.noaa.gov/mgg/sepm/palaios/9810/dimichele.html
http://www.museum.state.il.us/exhibits/mazon_creek/about_mazon_creek.html
http://www.creationinthecrossfire.com/documents/Varves/VarvesProblems.htm Thompson, Coelacanths, W. W. Norton & Company, New York and London, 1991 ISBN
0-393-02956-5, p. 78. Forey, P. 1998. History of the Coelacanth Fishes. Chapman
& Hall. Mary H. Schweitzer,* Mark Marshall, Keith
Carron, D. Scott Bohle, Scott C. Busse, Ernst V. Arnold, Darlene Barnard, J. R.
Horner*, and Jean R. Starkey, Heme compounds in dinosaur
trabecular bone, Proc. Natl. Acad. Sci. USA, Evolution, Vol. 94, pp.
6291-6296, June 1997 (
http://www.pnas.org/cgi/content/full/94/12/6291
) Debaun, J. (http://www.televar.com/~jnj/item6.htm)
- accessed Feb. 2004.
http://www.bio.davidson.edu/old_site/student/Lindthesis/Amy's%20Thesis.html
http://sprojects.mmi.mcgill.ca/immunology/spec_imm_cells.htm Tiffin, Bree, Bred in the bone: more clues about
the dinosaurs, Discovery Channel, EXN.ca, June 16, 1997 (
http://www.exn.ca/Stories/1997/06/12/03.asp
) Leonard Brand, Taphonomy of fossil whales in the
Miocene/Pliocene Pisco Fm., Peru, Dept. of Natural Sciences, Loma Linda
University, 2004 (
http://www.llu.edu/llu/grad/natsci/brand/whale.htm
) Stephen M. Rowland, THE ICHTHYOSAUR Nevada's State Fossil,
Heldref Publications, Nov 1999 ( Dr. Rowland is a professor of geology at the
University of Nevada, Las Vegas) (
http://articles.findarticles.com/p/articles/mi_m0GDX/is_6_74/ai_58243235
) Mary H. Schweitzer, Jennifer L.
Wittmeyer, John R. Horner, Jan B. Toporski, Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus
rex, Science, March 25, 2005 Edinger Collection (
http://www.lebendigevorwelt.de/megasucc/meg_13.en.htm
) Leonard Brand and James Florence, Stratigraphic
Distribution of Vertebrate Fossil Footprints Compared with Body Fossils -
Origins 9(2):67-74, 1982 ( http://www.grisda.org/origins/09067.htm
)
http://www.ldeo.columbia.edu/edu/dees/courses/v1001/images/horseshoecrab.gif John Horner, Digging Dinosaurs, 1988. (
http://www.ooblick.com/text/patton/
)
http://www.creationresearch.org/crsq/articles/32/32_4a1.html Baars, D.L., 1962. Permian System of the
Colorado Plateau. American Association of Petroleum Geologists Bulletin,
vol. 46, pp. 200-201. Middleton, L.T., Elliott, D.K. andMorales,
M., 1990. Coconino Sandstone. In: Grand Canyon Geology, S.S. Beus and M.
Morales (eds), Oxford University Press, New York, and Museum of Northern Arizona
Press, chapter 10, pp. 183-202. Young, D.A., 1990. The discovery
ofterrestrial history. In: Portraits of Creation, H.J. Van Till, R.E.
Shaw, J.H. Stek and D.A. Young (eds), William B. Eerdmans, Grand Rapids,
Michigan, chapter 3, pp. 80-81. McKee, E.D., 1947. Experiments on the
development of tracks in fine cross-bedded sand. Journal of Sedimentary
Petrology, vol. 17, pp. 23-28. Brand, L.R. and Tang, T., 1991. Fossil
vertebrate footprints in the Coconino Sandstone (Permian) of northern Arizona:
Evidence for underwater origin. Geology, vol. 19,pp. 1201-1204. Monastersky, R., 1992. Wading newts may
explain enigmatic tracks. Science News, vol. 141 (1), p. 5. Geology Today, vol. 8(3), May-June 1992,
pp, 78-79 (Wet tracks). Visher, G.S., 1990. Exploration
Stratigraphy, 2nd edition, Penn Well Publishing Co., Tulsa, Oklahoma, pp.
211-213. Kuenen, P.H. and Perdok, W.G., 1962.
Experimental abrasion: frosting and defrosting of quartz grains.
Journal of Geology, vol. 70, pp. 648-658. Amos, C.L. and King, E.L., 1984. Bedforms
of the Canadian eastern seaboard: a comparison with global occurrences.
Marine Geology, vol. 57, pp. 167-208. Brand, L.R. Origins
5(2):64-82 (1978). Brand, L.R. and T. Tang. 1991. Fossil
vertebrate footprints in the Coconino Sandstone [Permian] of northern Arizona:
evidence for underwater origin. Geology, 19:1201-1204. McKee, E.D. 1933. The Coconino Sandstone:
its history and origin. Carnegie Institution of Washington, Contributions to
Paleontology, Publication No. 440:77-115. McKee, E.D. 1944. Tracks that go uphill.
Plateau 16(4):61-73. McKee, E.D. 1945. Small-scale structures in
the Coconino Sandstone of northern Arizona. Journal of Geology 53(5):313-325.
http://www.studyworksonline.com/cda/content/article/0,,EXP888_NAV2-77_SAR879,00.shtml Lewin, R. Science, vol. 241, 15 July, p. 291 (1988) Raup, David M., Record is Catastrophic, Chicago Field
Museum of Natural History, Bulletin (Vol.54, March 1983), p.2 1, Univ. of
Chicago Dott Robert H., "The Rule", Presidential Address To
Society of Economic Paleontologists & Mineralogists, Geotimes, Nov. 1982, p.16 Pitman Sean D. Talk.Origins Debate (Link) Kendall Bioresearch Services (Link) Remembrance of A. K. Ghosh (Cambrian Gosh) by Satish
Srivastava, October, 2003 (Link) Michael A. Cremo, Paleobotanical Anomalies Bearing on the
Age of the Salt Range Formation of Pakistan: Ghosh, A. K, and Bose, A. (1947) Occurrence of microflora
in the Salt Pseudomorph Beds, Salt Range, Punjab. Nature, 160: 796-797. Ghosh, A. K., and Bose, A. (1950) Microfossils from the
Cambrian strata of the Salt Range, Punjab. Transactions of the Bose Research
Institute Calcutta, 18:71-78. Ghosh, A. K., and Bose, A. (1950b) Microfossils from the
Vindhyans. Science and Culture, 15: 330-331. Ghosh, A. K., and Bose, A. (1951) Recovery of vascular
flora from the Cambrian of Kashmir. Proceedings of the Indian Science
Congress, Part III, pp. 127-128. Ghosh, A. K., and Bose, A. (1951b) Evidence bearing on the
age of the Saline Series in the Salt Range of the Punjab. Geological Magazine,
88: 129-132. Ghosh, A. K., and Bose, A. (1952) Spores and tracheids
from the Cambrian of Kashmir. Nature, 169: 1056-1057. Ghosh, A. K., Sen, J., and Bose, A. (1948) Age of the
Saline Series in the Salt Range of the Punjab. Proceedings of the Indian
Science Congress, Part III, p. 145. Sahni, B. (1944) Age of the Saline Series in the Salt
Range of the Punjab. Nature, 153: 462-463. Sahni, B. (1945) Microfossils and problems of Salt Range
Geology. Proceedings of the National Academy of Sciences, India, Section B,
1944, Volume 14, pp. i-xxxii. Sahni, B. (1947a) The age of the Saline Series in the Salt
Range (Second Symposium).Concluding remarks. Proceedings of the National
Academy of Sciences, India, 1945, Section B, Volume 16, pp. 243-247. Sahni, B (1947b) Microfossils and the Salt Range Thrust.
Proceedings of the National Academy of Sciences, India, 1945, Section B,
Volume 16, pp. i-xlx. S. Leclercq, Evidence of Vascular Plants in the Cambrian,
Evolution, The International Journal of Organic Evolution
10: 109-114, June 1956 (Link) R. M. Stainforth, "Occurrence of Pollen and Spores in the
Roraima Formation of Venezuela and British Guiana," Nature, Vol. 210, No.
5033 (April 16, 1966), pp. 292--294. (Link) Jeanna Bryner, T. Rex
Related to Chickens, LiveScience (
Link
) Last Accessed: April 20, 2007; See also: Sharon Begley, T. Rex
and His Family, Newsweek, April 23, 2007 Hasiotis, ST, GSA Abstracts, A-461,
1997 Ariel Roth, Tom Zoutewelle, and
Dwight Hornbacher, Complex Concretions in the Jurassic Morrison Formation,
Geological Society of America, 2006 Abstracts With Programs 38(6):7 ( Link
) Dodson, P., A. K. Behrensmeyer, R. T. Bakker and J. S. McIntosh.
1980. Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison
Formation. Paleobiology 6(2):218 and 229.
Good, S., 1998, Bivalves as tools for paleoenvironmental
analysis--Upper Jurassic Morrison Formation of the Western Interior: final
report: in, C. Turner and F. Peterson, eds., Final Report: The
Morrison Formation Extinct Ecosystems Project, op. cit., pp.
121-158. Evanoff, E., Good, S., and Hanley, J., 1998, An overview
of the freshwater mollusks from the Morrison Formation (Upper Jurassic, Western
Interior, USA): Modern Geology, vol. 22, pp. 423-450. Turner, C., and Peterson, F., 1992, Sedimentology and
Stratigraphy of the Morrison Formation in Dinosaur National Monument, Utah and
Colorado: Annual Report of the National Park Service (unpublished),
contract #CA-1463-5-0001, in cooperation with the U.S. Geological Survey, 80 pp. Pierson, T., and Scott, K., 1985, Downstream dilution of a
lahar: transition from debris flow to hyperconcentrated streamflow: Water
Resources Research, vol. 21, pp. 1511-1524.
William A. Hoesch and Steven A. Austin, Dinosaur National
Monument: Jurassic Park or Jurassic Jumble? ICR, Impact Article # 370,
April, 2004 (Link)
Michael A. Cremo, Paleobotanical Anomalies Bearing on the Age of the Salt
Range Formation of Pakistan: A Historical Survey of an Unresolved Scientific
Controversy, Presentation at XXI International Congress of History of
Science, Mexico City, July 8-14, 2001 (Link)
Elizabeth A. Hadly and Brian A. Maurer, Spatial and temporal patterns of species
diversity in montane mammal communities of western North America,
Evolutionary Ecology Research, 2001, 3: 477-486 (
Link
)
Landing E, English A, Keppie JD.
Cambrian origin of all skeletalized metazoan phyla
discovery of Earth's oldest bryozoans (Upper
Cambrian, southern Mexico).
Geology 2010.38:547-550
Bernhart, Walter R., Dinosaur Nests Reinterpreted,
CRSQ Vol 41 No 2
September 2004 (
Link
) Johns, Warren H.,
Dinosaur eggs and
the post-Flood boundary, TJ 19(3) 2005 (
Link )
Tim Radford, Shell shock shows how dinosaurs suffered stress,
Science Editor,
The Guardian, Thursday
18 February 1999 22.55 EST (
Link
) . .
Home Page
. Truth, the
Scientific Method, and Evolution .
.
Maquiziliducks - The
Language of Evolution
. Defining Evolution
.
.
Evolving the
Irreducible
. . .
.
.
.
DNA Mutation Rates
. Donkeys, Horses,
Mules and Evolution .
.
.
Amino Acid
Racemization Dating
.The
Steppingstone Problem .
.
.
Harlen Bretz
. Milankovitch Cycles .
Kenneth Miller's Best
Arguments Since June 1, 2002  used to argue against transport. Observations in Spirit Lake
near Mount St. Helens and of trees uprooted by bulldozing operations show that
the small roots and rootlets are usually still intact, but the larger roots are
usually broken (See Figure). The presence of small roots extending from
the base of a petrified tree therefore is not evidence for an in situ
interpretation unless large roots also extend unbroken. Broken and frayed large
roots could be the result of changing stream currents eroding the bases of
growing trees, but such activity should leave evidences in the sediments.
Furthermore, erosion must be limited; otherwise, trees would be removed or
toppled.
used to argue against transport. Observations in Spirit Lake
near Mount St. Helens and of trees uprooted by bulldozing operations show that
the small roots and rootlets are usually still intact, but the larger roots are
usually broken (See Figure). The presence of small roots extending from
the base of a petrified tree therefore is not evidence for an in situ
interpretation unless large roots also extend unbroken. Broken and frayed large
roots could be the result of changing stream currents eroding the bases of
growing trees, but such activity should leave evidences in the sediments.
Furthermore, erosion must be limited; otherwise, trees would be removed or
toppled.
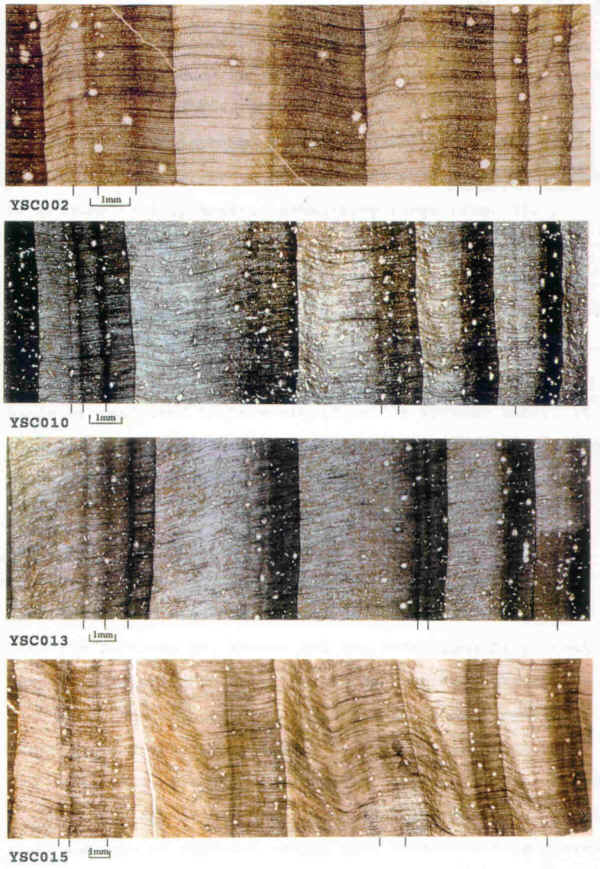 The flotation of organic matter (including trees in an upright
position) as illustrated in Spirit Lake at the base of Mount St. Helens provides
a model for interpreting the upright petrified trees of Yellowstone. Trees and
plants will float vertically when there is sufficient water and time. The
research at Spirit Lake helps explain the presence of the organic layer at the
level of the roots. Saturated organic debris sinks to the lake bottom to produce
a layer of organic matter. Upright floating trees also have dropped out of
suspension onto the bottom of Spirit Lake in a spacing pattern similar to that
of growing forests. They are not jumbled together in log jams as might be
expected. 20
The flotation of organic matter (including trees in an upright
position) as illustrated in Spirit Lake at the base of Mount St. Helens provides
a model for interpreting the upright petrified trees of Yellowstone. Trees and
plants will float vertically when there is sufficient water and time. The
research at Spirit Lake helps explain the presence of the organic layer at the
level of the roots. Saturated organic debris sinks to the lake bottom to produce
a layer of organic matter. Upright floating trees also have dropped out of
suspension onto the bottom of Spirit Lake in a spacing pattern similar to that
of growing forests. They are not jumbled together in log jams as might be
expected. 20 
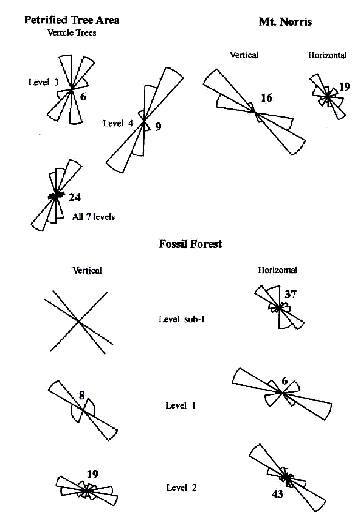 They lost all their roots, branches and bark, and were all lined up
in the same general direction, even the ones that sank into the lakes
vertically.
Fossilized trees can be found throughout the entire world and they all do
the same thing. Scientists at the National Petrified
Forest in Arizona freely speculate that the petrified trees of that forest were
washed into their current positions judging from the fact that they
generally have no branches, roots, or bark.3
They all appear as though they were in some sort of catastrophe
together. This is clearly visible to all who visit
such petrified forests.1
They lost all their roots, branches and bark, and were all lined up
in the same general direction, even the ones that sank into the lakes
vertically.
Fossilized trees can be found throughout the entire world and they all do
the same thing. Scientists at the National Petrified
Forest in Arizona freely speculate that the petrified trees of that forest were
washed into their current positions judging from the fact that they
generally have no branches, roots, or bark.3
They all appear as though they were in some sort of catastrophe
together. This is clearly visible to all who visit
such petrified forests.1  Yet another interesting finding concerning petrified trees, is that
many of them extend vertically through millions and millions of years of
sedimentary rock. How can this phenomenon be explained?
A common explanation is that these do not represent areas of the standard
geologic column, but areas of rapid local flooding and sedimentation.
Therefore, the layers that these trees
Yet another interesting finding concerning petrified trees, is that
many of them extend vertically through millions and millions of years of
sedimentary rock. How can this phenomenon be explained?
A common explanation is that these do not represent areas of the standard
geologic column, but areas of rapid local flooding and sedimentation.
Therefore, the layers that these trees  pass through do not represent thousands and millions of years.
However, the pictures shown to the right are of a petrified tree (located near
Katherine Hill Bay next to Flat Rocks Point, Australia) extending up
through many sedimentary layers and through two separated coal seams (See
Video Below).4
The tree itself is twelve feet tall, and was uncovered by a coal mining
company. If the two separated coal seams
represent long periods of time, how could this tree be
pass through do not represent thousands and millions of years.
However, the pictures shown to the right are of a petrified tree (located near
Katherine Hill Bay next to Flat Rocks Point, Australia) extending up
through many sedimentary layers and through two separated coal seams (See
Video Below).4
The tree itself is twelve feet tall, and was uncovered by a coal mining
company. If the two separated coal seams
represent long periods of time, how could this tree be  extending between them both? It
seems to me that this is a difficulty for the current understanding of science. Notice also that the layers themselves show no weathering
between one layer and the next even though each layer was supposedly the surface
of the earth for thousands if not millions of years. These combined
mysteries are more easily explained by rapid underwater burial with quickly
forming sediments. The theory that each fossil bearing layer in the
geologic column represents eons of time seems inadequate to explain such
problems that are easily explained by a quick catastrophic event.
extending between them both? It
seems to me that this is a difficulty for the current understanding of science. Notice also that the layers themselves show no weathering
between one layer and the next even though each layer was supposedly the surface
of the earth for thousands if not millions of years. These combined
mysteries are more easily explained by rapid underwater burial with quickly
forming sediments. The theory that each fossil bearing layer in the
geologic column represents eons of time seems inadequate to explain such
problems that are easily explained by a quick catastrophic event.  Coal itself is a problem. According to
modern science, coal forms from peat bogs.
Over a great many years, the peat becomes thicker and thicker.
After being covered by a sedimentary layer, the immense pressure and heat
combined with water, changes the organic material chemically into coal.
It is thought to take about 6 feet of organic material to make one foot
of coal. However, this theory has trouble explaining
a
Coal itself is a problem. According to
modern science, coal forms from peat bogs.
Over a great many years, the peat becomes thicker and thicker.
After being covered by a sedimentary layer, the immense pressure and heat
combined with water, changes the organic material chemically into coal.
It is thought to take about 6 feet of organic material to make one foot
of coal. However, this theory has trouble explaining
a 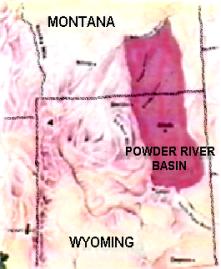 few interesting facts.
Coal seams, such as those found in the Powder River Basin of Gillette,
Wyoming, ranging upwards of 150 to 200 feet in thickness, point to a rapid
depositional and coalification process. Note the following comments published in
a 1993 article in Earth Magazine, "Powder River Coal: Geologic enigma,
environmental dilemma":
few interesting facts.
Coal seams, such as those found in the Powder River Basin of Gillette,
Wyoming, ranging upwards of 150 to 200 feet in thickness, point to a rapid
depositional and coalification process. Note the following comments published in
a 1993 article in Earth Magazine, "Powder River Coal: Geologic enigma,
environmental dilemma":
 Dinosaur tracks in coal
beds also pose another interesting problem.
Deep in the coalmines of southeastern Utah, literally thousands of
perfectly preserved Dinosaur footprints, of many different types, have been
found.
The imprint at the right is made through a clay layer and into the coal.
Dinosaur tracks in coal
beds also pose another interesting problem.
Deep in the coalmines of southeastern Utah, literally thousands of
perfectly preserved Dinosaur footprints, of many different types, have been
found.
The imprint at the right is made through a clay layer and into the coal.
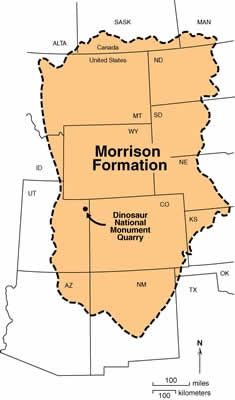 A further point of interest is that in some layers, on
occasion, a large species of dinosaur will be found in a particular layer with
almost nothing else in the same layer for these massive beasts to eat.1
What on earth did these dinosaurs live on for millions of years?
A further point of interest is that in some layers, on
occasion, a large species of dinosaur will be found in a particular layer with
almost nothing else in the same layer for these massive beasts to eat.1
What on earth did these dinosaurs live on for millions of years?  The Morrison is thought to have been a rather barren place with only
occasional oases in an overall vast scrubby or Savannah-like region. Yet, this
region preserved the remains of millions of very large plant-eating dinosaurs as
well as very large meat-eating dinosaurs.
For example, the dinosaur Apatosaurus is thought to have consumed
about 3 or 4 tons of vegetation on a daily basis.
The relative lack of preserved vegetation makes one wonder how such
massive beasts, millions of them, survived in this environment? (Link)
The Morrison is thought to have been a rather barren place with only
occasional oases in an overall vast scrubby or Savannah-like region. Yet, this
region preserved the remains of millions of very large plant-eating dinosaurs as
well as very large meat-eating dinosaurs.
For example, the dinosaur Apatosaurus is thought to have consumed
about 3 or 4 tons of vegetation on a daily basis.
The relative lack of preserved vegetation makes one wonder how such
massive beasts, millions of them, survived in this environment? (Link)













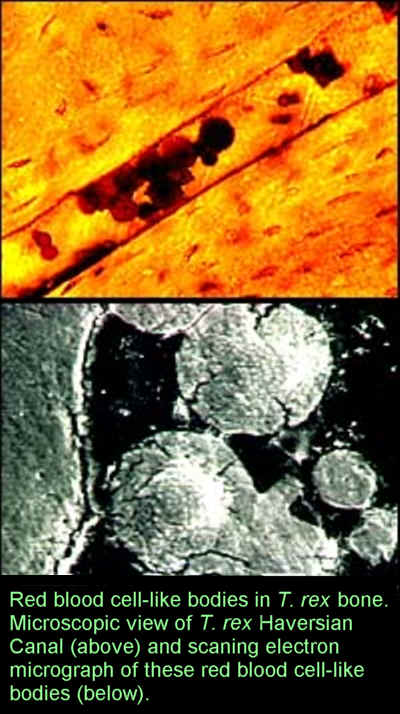 "The lab filled with murmurs of amazement, for I had focused on
something inside the vessels that none of us had ever noticed before: tiny round
objects, translucent red with a dark center. Then a colleague took one look at
them and shouted, 'You've got red blood cells. You've got red blood cells!'.
It was exactly like looking at a slice of modern bone. But, of course, I
couldn't believe it. I said to the lab technician: 'The bones, after all, are 65
million years old. How could blood cells survive that long?'"
10,11,50
"The lab filled with murmurs of amazement, for I had focused on
something inside the vessels that none of us had ever noticed before: tiny round
objects, translucent red with a dark center. Then a colleague took one look at
them and shouted, 'You've got red blood cells. You've got red blood cells!'.
It was exactly like looking at a slice of modern bone. But, of course, I
couldn't believe it. I said to the lab technician: 'The bones, after all, are 65
million years old. How could blood cells survive that long?'"
10,11,50
 These results are quite interesting since they indicate a very
specific immune response, not just against hemoglobin, but certain types of
hemoglobin molecules. Note again that the antibodies formed did not react
against snake hemoglobin indicating that the antibody reactivity was "specific
and not artifact." The question is, how much of the original T. rex
hemoglobin molecule would need to be intact to elicit such a specific immune
response in the laboratory rats?
These results are quite interesting since they indicate a very
specific immune response, not just against hemoglobin, but certain types of
hemoglobin molecules. Note again that the antibodies formed did not react
against snake hemoglobin indicating that the antibody reactivity was "specific
and not artifact." The question is, how much of the original T. rex
hemoglobin molecule would need to be intact to elicit such a specific immune
response in the laboratory rats?
 Now, it just seems quite unlikely that just 3 or 4 amino acids stuck
onto a heme group is going to give rise to an immune response at all not to
mention a specific immune response for a certain type of hemoglobin as was found
in this case (Note that a fully formed globin molecule ranges from 141 to
146 amino acids in length with specific folding characteristics that make up
various antigenic "epitopes" that antibodies detect). As
far as I have been able to tell, the degree of immune response specificity noted
by Schweitzer et al. has never been realized in any confirming experiment with
so few hemoglobin amino acids stuck to a heme group (just over 1,000da in size).
Even if such an experiment were successful, it certainly doesn't seem like a
very likely explanation in this case.
Now, it just seems quite unlikely that just 3 or 4 amino acids stuck
onto a heme group is going to give rise to an immune response at all not to
mention a specific immune response for a certain type of hemoglobin as was found
in this case (Note that a fully formed globin molecule ranges from 141 to
146 amino acids in length with specific folding characteristics that make up
various antigenic "epitopes" that antibodies detect). As
far as I have been able to tell, the degree of immune response specificity noted
by Schweitzer et al. has never been realized in any confirming experiment with
so few hemoglobin amino acids stuck to a heme group (just over 1,000da in size).
Even if such an experiment were successful, it certainly doesn't seem like a
very likely explanation in this case.  If this is not already enough, Schweitzer
recently made an even more startling discovery. About three years ago
(2002) she and her team had to divide a very large T. rex thigh bone in
order to transport it on a helicopter. When the bone was opened flexible soft
tissue "meat" was found inside. This is incredible because this bone was
supposed to be some 68 million
years old. Microscopic examination revealed fine delicate blood vessels with
what appear to be intact red blood cells and other type of cells like osteocytes
- which are bone forming cells. These vessels were still soft, translucent, and
flexible. Subsequent examination of other previously
excavated T. rex bones from this and other areas have also shown
non-fossilized soft tissue preservation in most instances.54
If this is not already enough, Schweitzer
recently made an even more startling discovery. About three years ago
(2002) she and her team had to divide a very large T. rex thigh bone in
order to transport it on a helicopter. When the bone was opened flexible soft
tissue "meat" was found inside. This is incredible because this bone was
supposed to be some 68 million
years old. Microscopic examination revealed fine delicate blood vessels with
what appear to be intact red blood cells and other type of cells like osteocytes
- which are bone forming cells. These vessels were still soft, translucent, and
flexible. Subsequent examination of other previously
excavated T. rex bones from this and other areas have also shown
non-fossilized soft tissue preservation in most instances.54
"I mean can you imagine pulling
a bone out the ground after 68 million years and then getting intact protein
sequences?" said John Asara of Beth Israel Deaconess Medical Center and Harvard Medical
School, lead author of one of the studies. "That's just mind boggling how much
preservation there is in these bones."
A Historical Survey of an Unresolved Scientific Controversy, For Presentation at
XXI International Congress of History of Science, Mexico City, July 8-14, 2001 (Link)
{kind=link}